Соответствующая корректура - Википедия - Conformational proofreading
Соответствующая корректура или же конформационный отбор это общий механизм молекулярное распознавание системы, в которых введение структурного несоответствия между молекулярным распознавателем и его мишенью или энергетическим барьером повышает специфичность и качество распознавания.[1][2][3][4][5][6] Проверка соответствия не требует энергозатрат и поэтому может использоваться в любой молекулярной системе распознавания. Соответствующая корректура особенно полезна в сценариях, где распознаватель должен выбрать подходящую цель среди множества похожих конкурентов.
Баланс между правильным и неправильным переплетом
Молекулярное распознавание происходит в шумной, переполненной биологической среде, и распознавателю часто приходится решать задачу выбора своей цели среди множества похожих конкурентов. Например, рибосома должна выбрать правильную тРНК, которая соответствует кодону мРНК среди многих структурно подобных тРНК. Если распознаватель и его правильная цель полностью совпадают, как замок и ключ, то вероятность связывания будет высокой, поскольку при связывании деформации не требуется. В то же время распознаватель может с большой вероятностью связываться с конкурентом с аналогичной структурой. Представляем структурное несоответствие между распознавателем (блокировкой) и ключом снижает вероятность привязки к правильной цели, но еще больше снижает вероятность привязки к аналогичной неправильной цели и, таким образом, улучшает специфичность. Тем не менее, слишком большая деформация резко снижает вероятность привязки к правильной цели. Таким образом, оптимальный баланс между максимальной вероятностью правильного связывания и минимизацией вероятности неправильного связывания достигается, когда распознаватель немного мимо. Это говорит о том, что конформационные изменения во время процессов молекулярного распознавания, такие как индуцированный припадок[7] Механизм, полезен для повышения специфичности распознавания.

Использование гомологичной рекомбинации для поиска гомологии
Механизм конформационная корректура используется в системе гомологичной рекомбинации для различения сходных последовательностей ДНК.[3][4] Гомологичная рекомбинация облегчает обмен генетическим материалом между гомологичными молекулами ДНК. Этот важный процесс требует обнаружения конкретной гомологичной последовательности ДНК в огромном разнообразии гетерологичных последовательностей. Обнаружение опосредовано RecA в E. coli или членами ее надсемейства в других организмах. RecA сначала полимеризуется вдоль отрезка одноцепочечной ДНК, а затем эта нить белок-ДНК ищет гомологию вдоль двухцепочечной ДНК. В филаменте RecA-ДНК расстояние между основаниями значительно увеличивается по сравнению с голыми 3,4 Å в двухцепочечной нити (в среднем на 50%).[8]). Это устанавливает значительный энергетический барьер при поиске, так как двухцепочечная ДНК должна растягиваться на такую же величину, чтобы проверить гомологию. Сформулировав процесс распознавания ДНК как проблему обнаружения сигнала, было показано, что экспериментально наблюдаемая деформация ДНК, вызванная RecA, и энергия связывания точно настроены для обеспечения оптимального обнаружения последовательности. Степень деформации такова, что связывание с гомологичными последовательностями ДНК уменьшается лишь незначительно, а связывание с неправильными последовательностями значительно уменьшается. Это как раз и есть механизм конформационной корректуры.
Экспериментальные доказательства конформационной корректуры путем гомологичной рекомбинации
Группа К. Деккера (Делфтский университет) непосредственно исследовала взаимодействия, участвующие в поиске гомологии, комбинируя магнитный и оптический пинцет.[9] Они обнаружили, что поиск и распознавание гомологии требует открытия спирали и, следовательно, может быть ускорен за счет раскручивания ДНК. Это в точности энергетический барьер, предсказываемый моделью конформационной корректуры. Данные указывают на физическую картину распознавания гомологии, в которой точность процесса поиска определяется расстоянием между сайтами связывания ДНК. Авторы приходят к выводу, что их интерпретация измерений «сродни конформационная корректура схема ... где дцДНК, а не нить RecA, является активным распознающим объектом поиска. Существует большое конформационное несоответствие между связанными с мишенью и несвязанными состояниями дцДНК. Доступ к связанному с целью состоянию осуществляется через энергетически невыгодные промежуточные состояния, как обсуждалось выше. Конформационное несоответствие улучшает селективность реакции узнавания ». Другими словами, они идентифицировали энергетический барьер и показали, что действительно двухцепочечная ДНК является активным участником, поскольку она должна преодолеть этот барьер.
Использование рибосомой для декодирования тРНК
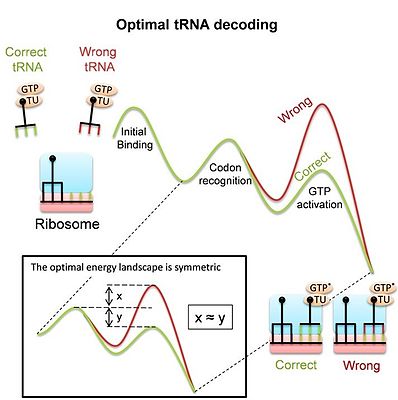
В рибосома представляет собой сложную молекулярную машину, которая для синтеза белков во время перевод процесс, должен декодировать мРНК путем объединения их кодонов с соответствующими тРНК. Декодирование является основным определяющим фактором приспособленности и требует точного и быстрого выбора правильных тРНК среди множества аналогичных конкурентов. Следует иметь в виду, что большинство событий связывания происходят из-за несовпадающих («несоответствующих») тРНК, и рибосома должна отвергать их как можно быстрее, чтобы освободить сайт связывания. В то же время рибосома должна удерживать соответствующие тРНК связанными достаточно времени, чтобы позволить процессу синтеза белка последовать. Несмотря на важность декодирования тРНК, до недавнего времени было неясно, действительно ли современная рибосома, и в частности ее большие конформационные изменения во время декодирования являются результатом адаптации к его задаче в качестве декодера или результатом других ограничений. Недавнее обучение[5]получили энергетический ландшафт, который обеспечивает оптимальное различение между конкурирующими субстратами тРНК и тем самым оптимальное декодирование тРНК. Оптимальный ландшафт - симметричный (см. Изображение). Исследование показывает, что измеренный ландшафт прокариотической рибосомы действительно симметричный. Эта модель предполагает, что конформационные изменения рибосомы и тРНК во время декодирования являются средством для получения такого оптимального декодера тРНК. Тот факт, что как гомологичная рекомбинация, так и декодирование тРНК используют конформационная корректура предполагает, что это общий механизм, который может широко использоваться системами молекулярного распознавания.
В других биологических системах
Ремонт УФ-повреждений человека
Недавнее исследование показывает, что конформационная корректура используется механизмами репарации ДНК человека.[10] Исследование было сосредоточено на вопросе о том, как белки репарации ДНК сканируют геном человека на наличие УФ -индуцированный урон на начальном этапе эксцизионная репарация нуклеотидов (NER). Детальные измерения одиночных молекул показали, как ДНК-связывающий белок, поврежденный ультрафиолетом (UV-DDB) выполняет поиск в 3D. Авторы обнаруживают, что «UV-DDB исследует сайты на ДНК дискретными шагами перед формированием долгоживущих, неподвижных димеров UV-DDB (DDB1 -DDB2 )2 в местах повреждения. Анализ скоростей диссоциации переходных связывающих молекул как на неповрежденной, так и на поврежденной ДНК показывает многократное время пребывания более трех порядков ... Считается, что эти промежуточные состояния представляют собой дискретные конформеры UV-DDB на пути к стабильному обнаружению повреждений » Авторы делают вывод из своих подробных кинетических измерений, что UV-DDB распознает очаги поражения, используя механизм проверки конформации через несколько промежуточных звеньев.
Другие схемы распознавания
Отношение к кинетической корректуре
в кинетическая корректура[11][12] схема, a временная задержка (эквивалентно, необратимая промежуточная стадия) вводится при образовании правильных или неправильных комплексов. Эта временная задержка снижает скорость продуцирования обоих комплексов, но повышает точность за пределами равновесного предела. Необратимость схемы требует источника энергии. Задержка при кинетической корректуре аналогична пространственной разнице при конформационной корректуре. Однако конформационная корректура может быть равновесной схемой, не потребляющей энергии.
Рекомендации
- ^ Савир Ю. и Тласти Т. (2007). Скалас, Энрико (ред.). «Конформационная корректура: влияние конформационных изменений на специфику молекулярного распознавания». PLoS ONE. 2 (5): e468. Bibcode:2007PLoSO ... 2..468S. Дои:10.1371 / journal.pone.0000468. ЧВК 1868595. PMID 17520027.
- ^ Савир Ю., Тласти Т. (2008). «Оптимальный дизайн молекулярного распознавателя: молекулярное распознавание как проблема обнаружения байесовского сигнала». Процесс передачи сигналов тем IEEE J Sel. 2 (3): 390–399. arXiv:1007.4527. Bibcode:2008ISTSP ... 2..390S. Дои:10.1109 / JSTSP.2008.923859.
- ^ а б c Савир Ю., Тласти Т. (2010). «RecA-опосредованный поиск гомологии как почти оптимальная система обнаружения сигналов». Молекулярная клетка. 40 (3): 388–96. arXiv:1011.4382. Дои:10.1016 / j.molcel.2010.10.020. PMID 21070965.
- ^ а б c Рэмбо Р.П., Уильямс Дж. Дж., Тайнер Дж. А. (2010). «Достижение точности гомологической рекомбинации, несмотря на чрезвычайную сложность: обоснованные решения с помощью молекулярного профилирования». Молекулярная клетка. 40 (3): 347–48. Дои:10.1016 / j.molcel.2010.10.032. ЧВК 3003302. PMID 21070960.
- ^ а б c Савир, Йонатан; Тлусты, Цви (11 апреля, 2013). «Рибосома как оптимальный декодер: урок молекулярного распознавания». Клетка. 153 (2): 471–9. Дои:10.1016 / j.cell.2013.03.032. PMID 23582332.
- ^ Алон У (2008). «Журнал клуб». Природа. 453 (7196): 701. Bibcode:2008Натура.453..701A. Дои:10.1038 / 453701e.
- ^ Кошланд Д. Э. (1958). «Применение теории ферментной специфичности к синтезу белков». Proc Natl Acad Sci U S A. 44 (2): 98–104. Bibcode:1958ПНАС ... 44 ... 98К. Дои:10.1073 / пнас.44.2.98. ЧВК 335371. PMID 16590179.
- ^ Чен З, Ян Х, Павлетич Н. П. (2008). «Механизм гомологичной рекомбинации из структур RecA-ssDNA / dsDNA». Природа. 453 (7194): 489–4. Bibcode:2008 Натур.453..489C. Дои:10.1038 / природа06971. PMID 18497818.
- ^ Де Вламинк I, ван Лоенхаут М.Т., Цвайфель Л., ден Бланкен Дж., Хунинг К., Хаге С., Керсемакерс Дж., Деккер С. (2012). «Механизм распознавания гомологии в рекомбинации ДНК из экспериментов с двумя молекулами». Молекулярная клетка. 46 (5): 616–624. Дои:10.1016 / j.molcel.2012.03.029. PMID 22560720.
- ^ Ghodke H, Wang H, Hsieh CL, Woldemeskel S, Watkins SC, Rapić-Otrin V, Van Houten B (6 мая 2014 г.). «Одномолекулярный анализ показывает, что человеческий ДНК-связывающий белок (УФ-DDB), поврежденный УФ-излучением, димеризуется на ДНК с помощью нескольких кинетических промежуточных продуктов». Proc Natl Acad Sci U S A. 111 (18): 1862–71. Bibcode:2014PNAS..111E1862G. Дои:10.1073 / pnas.1323856111. ЧВК 4020048. PMID 24760829.
- ^ Хопфилд Дж. Дж. (1974). «Кинетическая корректура: новый механизм уменьшения количества ошибок в биосинтетических процессах, требующих высокой специфичности». Proc Natl Acad Sci U S A. 71 (10): 4135–4139. Bibcode:1974PNAS ... 71.4135H. Дои:10.1073 / пнас.71.10.4135. ЧВК 434344. PMID 4530290.
- ^ Нинио Дж. (1975). «Кинетическое усиление дискриминации ферментов Biochimie». Биохимия. 57 (5): 587–595. Дои:10.1016 / S0300-9084 (75) 80139-8. PMID 1182215.