Специфическое для кишечника самонаведение - Gut-specific homing
Специфическое для кишечника самонаведение это механизм, с помощью которого активируется Т-клетки и клетки, секретирующие антитела (ASC), нацелены на оба воспаленный и невоспаленные области кишечника, чтобы обеспечить эффективное иммунная реакция. Этот процесс основан на ключевом взаимодействии между интегрин α4β7 и обращаться MadCAM-1 на поверхности соответствующих ячеек. Кроме того, это взаимодействие усиливается наличием CCR9, а хемокиновый рецептор, который взаимодействует с TECK. Витамин А -производный ретиноевая кислота регулирует экспрессию этих белков клеточной поверхности.
Резюме Т-клеток

Т-клетки производятся в вилочковая железа, а при выходе они мигрируют в лимфоидные органы тела, в том числе лимфатический узел. в паракортикальный слой лимфоидных узлов они подвергаются профессиональному антигенпрезентирующие клетки (БТР), например дендритные клетки (DC). Специфические взаимодействия между наивными Т-клетками и родственными им антигены приводит к активации Т-клеток. Активированные Т-клетки, иммунобласты, подвергаются клональной экспансии перед приобретением эффекторных функций. Затем активированные Т-клетки эмигрируют из лимфатических узлов через эфферентный лимфатический сосуд, и перемещаться по телу в кровь.[1]
При определенных обстоятельствах некоторые активированные Т-клетки предпочитают патрулировать определенные ткани. Это было названо возвращение лимфоцитов. Специфический для кишечника самонаведение - это преимущественное перемещение активированных Т-клеток к кишечник и кишка. Таким образом, Т-клетки эффективно задействуются, чтобы сформировать часть первой линии защиты от патогены. Это потому, что Т-клетки нацелены на первичные очаги инфекции и рециркулируют вокруг них. В целом это приводит к чрезвычайно высокой концентрации лимфоциты в этом регионе; 70% иммуноглобулин -продуцирующие клетки находятся в слизистая оболочка поверхности тела.[2]
История
Гризелли впервые предположил, что существуют разные пути миграции Т-клеток.[3]
Первоначальные теории о том, что Т-клетки нацелены не на ткань, а на антигены, были опровергнуты, так как работа Кэхилла показала, что очищенные Т-клетки перемещаются в область активации. Например, Т-клетки активированы в Патч Пейера лимфатические узлы, обнаруженные в кишечнике, вернутся в кишечник.[1] Последующие исследования пытались объяснить причину этого нацеливания на Т-клетки.

Первоначально предполагалось, что микросреда лимфоидных тканей является сигналом для преимущественного перемещения лимфоцитов.[4] Хотя до некоторой степени это может быть так, в настоящее время широко распространено мнение, что основной движущей силой самонаведения лимфоцитов является взаимодействие между Т-клетками. молекулы адгезии (рецепторы нахождения лимфоцитов ) и лиганды (адресины) на ткани венулы высокого эндотелия (HEV). Эта теория возникла из наблюдения, что интегрины клеточной поверхности различаются в разных популяциях Т-клеток. Начались работы по выявлению различий и их значения для миграции Т-клеток.
Стало очевидным, что уровни интегрина α4β7 были значительно выше на активированных Т-клетках по сравнению с наивными Т-клетками. Кроме того, этот интегрин гораздо чаще встречается на CD8 + Т-клетках, активированных в пейеровских бляшках (Т-клетки PP), по сравнению с другими подмножествами Т-клеток, такими как периферические и брыжеечные лимфатические узлы активированные Т-клетки.
Механизм
Механизм кишечного хоминга в значительной степени основан на взаимодействии между рецепторами.
Катящееся взаимодействие
Лимфоциты мигрируют из кровотока посредством характерного «перекатывающегося взаимодействия», опосредованного взаимодействиями между белками клеточной поверхности. Это «перекатывающееся взаимодействие» происходит в три этапа:
- Первоначальное связывание лимфоцита селектин и его лиганд. Это создает нестабильное взаимодействие, при котором лимфоцит легко удерживается эндотелий стена. Сила лимфы заставляет клетку характерно катиться по сосуду. Примером этого является взаимодействие между L-селектин и лиганд PNAD (адресация периферических узлов).
- Арест «катящейся» ячейки путем активации. Дальнейшие взаимодействия между белками или активация через хемокины вызывают формирование более стабильных сил между клеткой и сосудом. Теперь кювета прочно прикреплена к стенке сосуда. Продолжая пример, клетка будет заблокирована взаимодействием между ICAM и LFA-1 которые усилены хемокинами.
- Конформационное изменение. Активация клетки вызывает конформационное изменение, которое приводит к уплощению клетки относительно стенки сосуда. После сплющивания клетка может подвергнуться трансэндотелиальная миграция.[1]
Специфические белковые взаимодействия кишечника
«Самонаводящиеся рецепторы» имеют решающее значение в формировании взаимодействий, которые позволяют лимфоцитам двигаться.
Рецепторы вовлечены
MadCAM-1
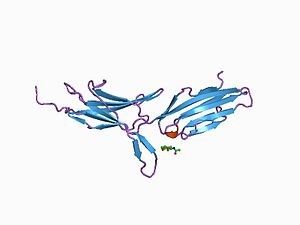
MadCAM-1 экспрессия непрерывна в венулах высокого эндотелия пейеровых бляшек и в микрососудах кишечного тракта.[5] Ингибиторы MadCAM-1 предотвращают миграцию Т-клеток в кишечник.
Структурный анализ белка MadCAM-1 показывает, что это трансмембранный белок 41,5 кДа с небольшим цитоплазматическим хвостом и большой внеклеточной областью. Эта внеклеточная область включает три иммуноглобиноподобных (Ig) домена. Последний на 33% структурно идентичен Са2 домен IgA-1. Предполагается, что первые два домена являются наиболее важными во взаимодействии α4β7.
Белок MadCAM-1 имеет структурную гомологию с рецепторами сосудистой поверхности VCAM1 и ICAM на 28% и 32% соответственно.[6]
Интегрин Α4β7
Это выражается на активированных Т-клетках, импринтированных для специфического для кишечника самонаведения. Это контррецептор для белка MadCAM-1, взаимодействия которого вызывают начальный этап взаимодействия в модели «катящихся» клеток миграции лимфоцитов.
CCR9
CCR9 рецепторы представлены на Т-клетках и участвуют в формировании более стабильного взаимодействия между лимфоцитом и стенкой сосуда. Лигандом для белков CCR9 является CCL25 хемокин (также известный как TECK) белок. CCL25 представляет собой хемокин, который при связывании со своим рецептором CCR9 на лимфоците активирует конформационное изменение в поверхностных интегринах этого лимфоцита,[7] обеспечение высокоаффинного связывания со стенкой сосуда и последующего переселение.
Хемокин CCL25 преобладает в эпителиальной ткани тонкой кишки (7). Было показано, что CD8 + Т-клетки, активированные в лимфатической ткани пейерова пятна, проявляют тропизм в сторону хемокина CCL25. Т-клетки мигрируют вверх по градиенту к CCL25.[4]
Другие клетки
Все GALT (лимфоидная ткань, ассоциированная с кишечником ) DC, с α-цепочкой CD103, способны продуцировать Т-клетки, возвращающиеся в кишечник. Исходя из этого, было высказано предположение, что дендритные клетки каким-то образом импринтируются, прежде чем достигнут лимфоидных органов, чтобы вызвать способность самонаведения.[8]
Регулирование витамина А
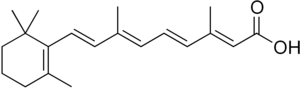
Витамин А играет фундаментальную роль в ответной реакции кишечника. Данные показывают, что витамин А превращается в ретиноевая кислота (RA) путем окисления в дендритных клетках пейерсовских пятен. Присутствие RA приводит к экспрессии ключевых рецепторов хоминга, α4β7 и CCR9, на клеточной поверхности Т-клеток, потенциально посредством регуляции транскрипции.
Производство ретиноевой кислоты
Алкогольдегидрогеназы (ADH) и ретинальная дегидрогеназа (RALDH) ферменты катализируют окисление пищевого витамина А до ретиноевой кислоты. Именно присутствие этой ретиноевой кислоты индуцирует экспрессию CCR9 и α4β7.[9] Более низкие концентрации ретиноевой кислоты внутри клетки приводят к снижению экспрессии хемокинового рецептора CCR9, что, в свою очередь, ограничивает способность клетки проникать в слизистую оболочку кишечника. Хотя экспрессия α4β7 также снижается при более низких концентрациях ретиноевой кислоты, предполагается, что уровни ретиноевой кислоты должны быть истощены в большей степени, чтобы повлиять на экспрессию α4β7 по сравнению с CCR9.[10]
Вестерн-блоттинг показал, что дендритные клетки из участков Пейера продуцируют как белки RALDH1, так и ADH. Кроме того, было обнаружено, что они экспрессируются в эпителиальных клетках тонкого кишечника. Следовательно, эти клетки способны превращать витамин А в основную физиологическую форму RA: полностью транс-RA. Исследования, демонстрирующие способность ДК катализировать превращение ретинола в ретиноевую кислоту, подтверждают это предположение.[10]

Влияние на экспрессию рецепторов
Исследования на мышах показали, что уровень ретиноевой кислоты в клетках действительно оказывает значительное влияние на экспрессию рецепторов на поверхности клетки. Модели на мышах показали, что даже увеличение концентрации ретиноевой кислоты в клетках на 0,1 нМ увеличивает экспрессию α4. Кроме того, экспрессия β7 увеличивалась за счет повышения концентрации клеточной ретиноевой кислоты. Чтобы гарантировать, что увеличение концентрации RA происходит за счет каталитических ферментов ADH и RALDH, использовали ингибиторы этих ферментов. Цитраль, ингибитор фермента RALDH, подавлял экспрессию адгезинов α4β7 на поверхности клетки, что свидетельствует о важности катализа витамин A → ретиноевой кислоты.
Способность самонаведения
Это, теоретически, должно увеличить способность активированных Т-клеток к кишечнику. Доказательства этого были получены в исследованиях с участием мышей, получавших лечение РА и мышей с дефицитом витамина А. У мышей с повышенным уровнем RA в ткани кишечника было обнаружено большее количество Т-клеток. Исследования на мышах с дефицитом витамина А подтвердили, что верно обратное, поскольку в кишечнике обнаружено значительно меньшее количество Т-клеток. собственная пластинка по сравнению с контрольными мышами. Напротив, такого снижения количества Т-клеток в легких и печени дефицитных мышей не наблюдалось, что означает, что путь импринтинга RA уникален для кишечно-специфических самонаводящихся Т-клеток.[11]
Регуляция транскрипции ретиноевой кислотой
Было высказано предположение, что уровни RA влияют на экспрессию рецепторов MadCAM-1 и α4β7 посредством регуляции транскрипции.
Рецепторы RAR и RXR, которые являются нуклеиновыми рецепторами для RA, могут образовывать гетеродимеры при определенных клеточных условиях. В конформации димера рецепторы способны связывать последовательности RARE или RXRE, обнаруженные в ДНК. Эти генетические области часто находятся на 5'-конце генов, что позволяет предположить, что они играют регулирующую роль. Фактически, они часто находятся в начале или в интронах генов, регулируемых в ответ на уровни ретиноидов.
Участки этих последовательностей RARE, а также RA-боксы были обнаружены в первых экзонах генов, кодирующих b7 и рецепторы CCR9. Использование белков-антагонистов против субъединицы RAR показало снижение экспрессии альфа.[12]
Следовательно, возможно, что экспрессия альфа-рецепторов транскрипционно регулируется концентрацией RA в клетке.
Другие эффекты витамина А на возвращение лимфоцитов
Было также показано, что повышенные концентрации витамина А снижают экспрессию рецепторов P-Lig, E-Lig и Fuct-VII in vitro. Это говорит о том, что витамин А не только увеличивает способность Т-клетки к проникновению в кишечник, но и одновременно снижает вероятность связывания этой клетки с другой тканью.
Причины самонаведения лимфоцитов
Самый убедительный аргумент заключается в том, что, направляя лимфоциты в область, в которой они были активированы, более вероятно, что они войдут в контакт со своим родственным антигеном. Они нацелены на область, где антиген патогена наиболее вероятно находится на периферии. Кроме того, возможно, что тип ткани может влиять на фенотипический свойства Т-клеток как при активации, так и при миграции через ткань. Поэтому казалось бы разумным, чтобы Т-клетки проявляли признаки предпочтительной ткани, поскольку они, вероятно, будут выполнять свою защитную роль в среде, для которой они наиболее подходят.
Проблемы со здоровьем
У пожилых людей часто наблюдается возрастное снижение иммунного ответа кишечника. Эта иммунное старение была выдвинута гипотеза, что это связано с нарушением самонаведения лимфоцитов к пейерсовским пятнам, обнаруженным в кишечнике.[13]
Этот процесс не уникален для людей и задокументирован у других животных, включая макак-резус. [14] и крысы. Исследования показали, что у старых крыс снижена экспрессия ключевых кишечных рецепторов α4β7; и MadCAM-1. У этих старых крыс было на 30% меньше α4β7; и на 17% меньше MadCAM-1, чем его эквиваленты у молодых крыс.[15]
Хотя на сегодняшний день причинно-следственная связь не доказана, предполагается, что снижение экспрессии рецепторов отвечает за общую потерю иммунного ответа. Кроме того, посттрансляционные модификации могут ослаблять сродство рецепторов друг к другу.[15]
Будущая работа
Требуется гораздо большая работа, чтобы полностью понять условия, необходимые для возникновения тканеспецифической самонаведения. Однако при более глубоком понимании этой области открывается большой потенциал в области медицинской терапии. Может стать возможным нацеливание на оральный аттенуированные вакцины к слизистой оболочке кишечника. Мы надеемся, что таким образом вакцина станет более надежной и эффективной. Кроме того, он может улучшить лечение хронических заболеваний кишечника.
Направление других тканеспецифичных лимфоцитов
Специфический хоминг в кишечнике является наиболее понятным из хоминга лимфоцитов. Однако есть много других примеров, в том числе:
- Лимфоидное специфическое самонаведение- Возвращение наивных Т-клеток в лимфатическую ткань по всему телу. Адгезин, экспрессируемый наивными Т-клетками, представляет собой L-селектин (1). как только Т-клетки активируются, они подавляют экспрессию L-селектина, позволяя им свободно перемещаться по телу в крови.
- Самонаведение для кожи- Действует как метод набора большого количества Т-клеток памяти в кожу, что создает высокий риск инфицирования. Опять же, это включает экспрессию различных интегринов. В этом случае у человека α4β1 и кожный лимфоцитарный антиген (CLA ) экспрессируются на поверхности Т-клеток. Эти интегрины взаимодействуют с E-selectin молекулы, обнаруженные на воспаленной коже.
- Грудное самонаведение- Участвует в перенаправлении Т-клеток в молочные железы. это происходит для того, чтобы защитные лимфоциты попадали в грудное молоко.
- Самонаведение В-клеток- В последнее время об этом мало что известно.
использованная литература
- ^ а б c Маккей (1993). «Возвращение наивных, память и эффекторных лимфоцитов». Текущее мнение в иммунологии. 5 (3): 423–427. Дои:10.1016 / 0952-7915 (93) 90063-Х. PMID 8347302.
- ^ фон Андриан, Мемпель (2003). «Самонаведение и сотовый трафик в лимфатических узлах». Nature Reviews Иммунология. 3 (11): 867–878. Дои:10.1038 / nri1222. PMID 14668803.
- ^ Ивата, Хиракияма; и другие. (2004). «Ретиноевая кислота накладывает отпечаток специфичности на Т-лимфоциты по отношению к кишечнику». Журнал клинических исследований. 21 (8): 527–538. Дои:10.1172 / JCI44262. ЧВК 3223921. PMID 21737878.
- ^ а б Родриго Мора, Ноно, Манджунатх; и другие. (2003). «Селективный импринтинг Т-клеток, возвращающихся в кишечник, дендритными клетками Пейера». Природа. 424 (6944): 88–93. Bibcode:2003Натура.424 ... 88М. Дои:10.1038 / природа01726. PMID 12840763.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Шипперс, А; Leuker, C; Пабст, О; Кочут, А; Прохнов, Б; Gruber, AD; Leung, E; Криссансен, GW; Вагнер, Н; Мюллер, В. (сентябрь 2009 г.). «Молекула адгезии клеток адгезии слизистой оболочки-1 контролирует миграцию и функцию плазматических клеток в тонком кишечнике мышей». Гастроэнтерология. 137 (3): 924–33. Дои:10.1053 / j.gastro.2009.05.039. PMID 19450594.
- ^ Брискин; и другие. (1996). «Структурные требования для связывания адресаина слизистой оболочки с его рецептором лимфоцитов альфа 4 бета 7. Общие темы среди взаимодействий семейства интегрин-Ig». Журнал иммунологии. 156 (2): 719–726. PMID 8543825.
- ^ Пармо-Кабаньяс, М; Гарсия-Берналь Д.; Гарсия-Вердуго Р; Кремер Л; Márquez G; Тейксидо Дж. (Август 2007 г.). «Внутриклеточная передача сигналов, необходимая для CCL25-стимулированной адгезии Т-клеток, опосредованной интегрином альфа4бета1». J. Leukoc. Биол. 82 (2): 380–91. Дои:10.1189 / jlb.1206726. HDL:10261/72227. PMID 17510295.
- ^ Йоханссо-Линдбом; и другие. (2005). «Функциональная специализация CD103 + дендритных клеток кишечника в регуляции селективного хоминга Т-клеток». Журнал экспериментальной медицины. 234 (1): 268–281. Дои:10.1111 / j.0105-2896.2009.00874.x. PMID 20193025.
- ^ Дестер (2000). «Семейства ретиноиддегидрогеназ, регулирующих функцию витамина А: производство зрительного пигмента и ретиноевой кислоты». Европейский журнал биохимии. 8 (14): 4315–24. Дои:10.1046 / j.1432-1327.2000.01497.x. PMID 10880953.
- ^ а б Берлин (1993). «Интегрин альфа4бета7 опосредует связывание лимфокса с сосудистым адресом слизистой оболочки MAdCAM-1». Ячейка. 74 (1): 185–195. Дои:10.1016 / 0092-8674 (93) 90305-а. PMID 7687523.
- ^ Верснель (2004). «Ретиноевая кислота: обучающий« витаминный эликсир »для кишечных Т-лимфоцитов». Иммунитет. 21 (4): 458–460. Дои:10.1016 / j.immuni.2004.10.002. PMID 15485623.
- ^ Ивата; и другие. (2004). «Ретиноевая кислота отпечатывается на Т-лимфоцитах. Иммунитет. 21 (4): 527–538. Дои:10.1016 / j.immuni.2004.08.011. PMID 15485630.
- ^ Schmucker, DL; Оуэн, Р.Л .; Outenreath, R; Торо, К. (2003). «Основа возрастного снижения иммунитета слизистой оболочки кишечника». Клиническая иммунология и иммунология развития. 10 (2–4): 167–72. Дои:10.1080/10446670310001642168. ЧВК 2485420. PMID 14768948.
- ^ Тейлор, LD; Дэниелс, СК; Шмукер, DL (апрель 1992 г.). «Старение снижает иммунный ответ слизистой оболочки желудочно-кишечного тракта у макак-резусов». Иммунология. 75 (4): 614–8. ЧВК 1384839. PMID 1592437.
- ^ а б Schmucker, Douglas L; Оуэн, Тревор М; Issekutz, Thomas B; Гонсалес, Лоида; Ван, Роза К. (август 2002 г.). «Экспрессия рецепторов самонаведения лимфоцитов α4β7 и MAdCAM-1 у молодых и старых крыс». Экспериментальная геронтология. 37 (8–9): 1089–1095. Дои:10.1016 / S0531-5565 (02) 00065-7. PMID 12213559.