Макроолит - Википедия - Macroolithus
| Макроолитус | |
|---|---|
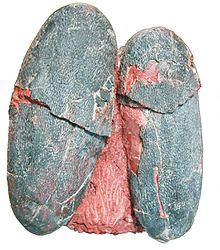 | |
| Пара Macroolithus yaotunensis яйца | |
| Классификация окаменелостей яиц | |
| Базовый тип оболочки: | Орнитоз |
| Морфотип: | Орнитоид-ратит |
| Семья: | †Elongatoolithidae |
| Оогенус: | †Макроолитус Чжао, 1975 г. |
| Тип oospecies | |
| †Оолит ругуст Молодые, 1965 г. | |
| Виды | |
| |
Макроолитус является ооген (род ископаемых яиц) яйцо динозавра принадлежащий к семья Elongatoolithidae. В типовой вид, М. rugustus, первоначально был описан под ныне несуществующим названием oogenus Оолиты. Известны еще три вида: М. yaotunensis, М. mutabilis, и М. lashuyuanensis. Это относительно крупные яйца удлиненной формы с двухслойной скорлупой. Их гнезда состоят из больших концентрических колец парных яиц. Есть свидетельства сине-зеленой пигментации его панциря, которая, возможно, помогала замаскировать гнезда.
Макроолитус были обнаружены яйца, содержащие овирапторид динозавр эмбрионы напоминающий Heyuannia. Множественные другие ассоциации между овирапторидом и элонгатоолитидом (включая другие яйца, содержащие эмбрионы, родители, вынашивающие яйца в гнездах, и пара очищенных от скорлупы) Макроолитус-подобные яйца, сохранившиеся в тазу овирапторида) подтверждают, что родитель Макроолитус был овирапторидом.
Он находится в Верхний мел образования Центральной и Восточной Азии; окаменелости были найдены в Монголии, Казахстане и Китае. В формации Наньсюн в Южном Китае Макроолитус окаменелости варьируются до и, возможно, более Граница мелового и третичного периода, который традиционно считается знаком исчезновенияптичий динозавры. Некоторые палеонтологи интерпретировали появление яиц динозавров в этом образовании как свидетельство постепенного вымирания, а не внезапного катастрофического события. Однако другие палеонтологи полагают, что эти интерпретации просто основаны на артефактах эрозии и переотложения в ранние годы. Палеоген.
Описание

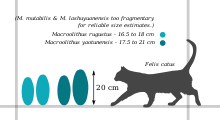
Макроолитус Яйца характеризуются большим размером, от 16 до 21 см (от 6,3 до 8,3 дюйма) в длину, и особенно грубым орнаментом.[1][2] Их микроструктура недостаточно хорошо описана в литературе,[1] но обычно следует типичному образцу элонгатоолитида:[2] Яичная скорлупа разделена на два структурных слоя (маммиллярный слой и сплошной слой). Сплошной слой образует внешнюю часть яичной скорлупы; его единицы яичной скорлупы сплавлены вместе, так что слой кажется сплошным. Внутренний слой, известный как маммиллярный или конический слой, состоит из конусовидных структур, образующих основу яичной скорлупы.[1] В Макроолитуссплошной слой в два-три раза толще мамиллярного слоя.[1][2] Яйца имеют большое разнообразие пористой структуры. Поскольку газовая проводимость связана с размером пор и плотностью, это может означать, что яйца были отложены в различных условиях.[1]
Виды
Четыре вида Макроолитус в настоящее время признаются:
- М. rugustus - в типовой вид, эти яйца находятся в Nanxiong и Цзянси на юге Китая; это также распространено в Формация Немегт из Монголия и Манракская Свита в Казахстан.[3][4] Его размер варьируется от 16,5 до 18 см (от 6,5 до 7,1 дюйма) в длину и от 7,5 до 8,5 см (от 3,0 до 3,3 дюйма) в поперечнике. Его яичная скорлупа имеет толщину от 0,8 до 1,7 мм (от 0,031 до 0,067 дюйма), из которых около 1/4 составляет маммиллярный слой.[5] Наружная поверхность покрыта неправильными цепочками узлов (бугорчатая скульптура) вокруг внешнего центра яйца, переходящими в более ямчатый, сетчатый узор (сагенотуберкулезный) на концах.[6][7]
- М. yaotunensis является родным для Хэнань и чтобы Nanxiong.[8][5] Имеет микроструктуру и орнамент, аналогичные М. rugustus, но их можно отличить по тому, что они немного больше (от 17,5 до 21 см (от 6,9 до 8,3 дюйма) в длину) и имеют более толстую оболочку (от 1,4 до 1,9 мм (от 0,055 до 0,075 дюйма)).[1][2] Также в отличие от М. rugustus, М. yaotunensis показывает волнистую границу между его маммиллярным слоем и его сплошным слоем.[5]
- М. mutabilis - этот монгольский вид известен по нескольким фрагментам яичной скорлупы. Фрагменты имеют толщину от 1,3 до 2,0 мм (от 0,051 до 0,079 дюйма), что немного толще, чем М. rugustus. В собранном виде размер яйца, вероятно, превышает 17 см (6,7 дюйма). Он уникален тем, что Михайлов назвал «аберрационными» особенностями: некоторые фрагменты имеют щелевидные поровые каналы и лишены орнаментации.[2]
- М. lashuyuanensis - этот вид основан на фрагменте яичной скорлупы, найденном в Бассейн Наньсюн из Гуандун. Его яичная скорлупа толщиной от 2,3 до 2,7 мм (от 0,091 до 0,106 дюйма) является самой толстой среди Макроолитус виды. Кроме того, он отличается наличием темной плотной полосы в верхней части непрерывного слоя и постепенным переходом между непрерывным и маммилярным слоями.[9]
Окраска
Некоторые образцы М. yaotunensis сохранить следы красновато-коричневого пигмента протопорфирин и сине-зеленый пигмент биливердин, те же пигменты, которые используются в яйцах многих современных птиц. Скорее всего, яйца были интенсивного сине-зеленого цвета, о чем свидетельствует преобладание биливердина в скорлупе. Цель окраски неясна, но у современных птиц она может служить окраской или послеродовым сексуальным сигналом. Цветные яйца также связаны с активной отцовской заботой у современных птиц, что еще раз подтверждает гипотезу о том, что самцы овирапторид заботятся о своих яйцах.[10]
Имена и этимология
Название Макроолитус происходит от греческих корней макро- (что означает "большой")[11] и оолит (что означает «каменное яйцо»), обычный суффикс для названий оогенусов.[12] Его китайское название (巨 形 蛋 Jùxíng dàn) аналогично переводится как «гигантское яйцо».[5] Ооспецифические эпифеты yaotunensis (Китайский : 窑 屯 yáotún) и lashuyuanensis (Китайский : 腊 树 园 Làshùyuán) чтят места, где эти виды были первоначально обнаружены (яотун[5] и Лашуюань[9] районов Гуандуна соответственно), добавив латинский суффикс -ensis для обозначения места происхождения товара;[13] название mutabilis на латыни означает изменчивый.[2] Когда он назвал "Оолитс" ругуст, Ян (1965) не дал этимологии ругуст, но китайская форма (粗 皮 cūpí) переводится как «грубая кожа».[3]
Палеобиология и воспитание

Многие ассоциации между скелетами или эмбрионами взрослых овирапторозавров с яйцами элонгатоолитид (включая Макроолитус) демонстрируют, что Макроолитус и другие элонгатоолиты были заложены овирапторозаврами.[1][14] Один скелет овирапторозавра из Верхний мел из Китай был описан в 2005 году; внутри таза сохранились два яйца элонгатоолитид в скорлупе. Это говорит о том, что овирапторозавры имели два функциональных яйцеводы где оба производят яйца одновременно. Пока Сато и другие. не относил яйца специально к Макроолитус, они отметили, что яйца очень напоминают М. yaotunensis, правда, с более тонкой яичной скорлупой. Однако тонкая яичная скорлупа могла быть просто результатом того, что скорлупа еще не закончила формирование, когда мать умерла, или из-за биохимического растворения скорлупы до окаменения.[15]
Множественные разные роды овирапторид были обнаружены на гнездах элонгатоолитов или рядом с ними, что указывает на то, что родители овирапторид будут выводок на свои яйца, скорее всего, в течение длительного периода времени.[16] Есть некоторые свидетельства того, что овирапторид и троодонтид о яйцах заботился отец, возможно, в полигамный система.[17][10] Учитывая большой размер яиц по сравнению с родительским, мать откладывала бы только два яйца за раз, поэтому яйца одного гнезда могли быть внесены несколькими самками.[15][17]
Гнезда

Множественные хорошо сохранившиеся Макроолитус известны гнезда, представляющие М. rugustus и М. yaotunensis.[3][5] Очень хорошо сохранившаяся кладка М. yaotunensis содержит 20 яиц, расположенных в два, возможно, три круглых слоя. По завершении это гнездо могло содержать 40 или более яиц.[3]
Танака и другие. (2015) обнаружили, что у современных архозавров (группа, включающая птиц, крокодилов и динозавров) пористость яичной скорлупы может использоваться для точного прогнозирования того, закопаны ли яйца или отложены в открытых гнездах. Они пришли к выводу, что Макроолитус гнезда были либо открыты, либо частично покрыты растительностью.[18] Сине-зеленая окраска яиц могла служить камуфляжем, скрывающим гнездо от хищников.[10]
М. rugustus яйца в Цаган Хушу в Монголии представляют собой прибрежное колониальное гнездовье.[2]
Эмбрионы
Немного Макроолитус яйца сохраняют внутри эмбриональные остатки овирапторид.[19] Два яйца с эмбрионами, обнаруженные в Верхний мел Формация Наньсюн возле Ганьчжоу, Цзянси были упомянуты М. yaotunensis в 2008 году. Один из этих эмбрионов показывает гораздо большую степень развития костей (окостенение ), чем другой; сохраняет окостеневшие задние конечности и несколько позвонков. Дело в том, что шейные позвонки не только окостенели центр и нервные дуги, но и окостенели зигапофизы, привел Ченг и другие. сделать вывод, что этот эмбрион на самом деле может представлять детеныша, поскольку в современном куры и поморники эти зигафизы до вылупления являются хрящевыми. Также анатомия стопы эмбриона (в частности, пропорции ее плюсневые кости ) напоминает овирапторид Heyuannia huangi, что указывает на принадлежность этих яиц к Х. хуанги или аналогичный вид. Другое яйцо имеет гораздо менее развитый зародыш, сохранились только задние конечности. Хотя это не позволяет проводить сравнение на уровне видов, большеберцовая кость (большеберцовая кость) подтверждает, что это овирапторозавр.[19]

Еще три яйца, содержащие эмбрионы из той же формации, были описаны в 2016 г. и другие. Хотя они отметили, что эти яйца сильно напоминают М. yaotunensis, они отказались относить их к какому-либо оотаксону ниже Elongatoolithidae, потому что Макроолитус четко не определен и требует доработки. Эмбрионы в этих яйцах являются одними из наиболее хорошо сохранившихся овирапторид, что дает новую информацию об онтогенезе овирапторид. Образцы демонстрируют относительно неглубокую голову, что указывает на то, что по мере созревания овирапторид их черепа росли дорсовентрально (сверху вниз) быстрее, чем в переднезаднем направлении (спереди назад). Такая модель роста необычна среди теропод, но также наблюдается у полученный ("передовой") тираннозавриды. Также необычно то, что даже на этой ранней стадии разработки носовые кости слиты. По совпадению, тираннозавриды также демонстрируют слияние носовых костей на ранних стадиях развития.[14]
Патология и угасание
Много Макроолитус экземпляры из Южного Китая имеют двух- или многослойные шишки на внутренней поверхности яичной скорлупы, патологическое состояние известная как яйцеклетка в яйце.[20] Особенно распространен среди яиц, ближайших к Граница мелово-третичного периода (K-T), который представляет собой конец Мезозойская эра. Эта патология коррелирует с более высокой концентрацией микроэлементы подобно Co, Cr, Cu, Mn, Ni, Pb, Sr, V, и Zn.[20] Эксперименты на современных птицах показали, что воздействие высоких уровней этих элементов заставляет их включаться в яичную скорлупу, но точный механизм, лежащий в основе патологического многослойного образования, неизвестен. Эти аномалии предположительно повлияли на выводимость яиц и, возможно, сыграли роль в исчезновении динозавров.[14][20] Однако зародыш остается внутри трех многослойных Макроолитус-подобные яйца из Ганьчжоу кажутся нетронутыми.[14]
Граница K-T связана с аномально высокое количество иридия, элемент, который редко встречается в земной коре, но относительно часто встречается в астероидах и в ядре Земли. Это было использовано в качестве доказательства того, что удар метеорита вызвал вымирание динозавров ( Гипотеза Альвареса ). Однако некоторые палеонтологи связывают событие вымирания и аномалию иридия с более постепенным изменением климата, вызванным вулканической активностью Деканские ловушки. Бассейн Наньсюн имеет особое отношение к этому обсуждению, поскольку он содержит границу К-Т. Однако положение границы, а также продолжительность исчезновения были предметом споров. Чжао и другие. (2002 и 2009) постулировали, что произошло по крайней мере два события с доставкой иридия в течение периода времени, согласующиеся с вулканизмом Декканских ловушек, что коррелировало с постепенным сокращением разнообразия и, в конечном итоге, исчезновением окаменелых яиц из бассейна Наньсюн. Согласно их интерпретации, Макроолитус пережил 250 000 лет до начала Палеоцен.[21][22][23] Однако Бак и другие. (2004) оспаривали эти утверждения, утверждая, что селевые потоки смешал и переработал отложения вблизи границы, вызывая размытие иридиевой аномалии и появление фрагментов яичной скорлупы с обеих сторон.[24]
Классификация
Ископаемые яйца классифицируются по собственной паратаксономической системе, параллельной Линнеевская таксономия. Макроолитус, Elongatoolithus, и Nanhsiungoolithus были первыми оогенерами, когда-либо названными в этой системе; они были классифицированы в ofamily Elongatoolithidae.[5] Кладистический анализ также поддерживает размещение Макроолитус рядом с Elongatoolithusвместе с другими элонгатоолитами в клады.[25][26] Известно четыре вида: М. rugustus, М. yaotunensis, М. mutabilis, и М. lashuyuanensis.[1][27] Микроструктура оогенуса плохо выражена.[1] и поэтому может нуждаться в доработке.[14] Когда Михайлов описал М. mutabilis, он не обнаружил микроструктурных различий в оригинальных иллюстрациях Чжао М. rugustus и М. yaotunensis, но не подбирать синонимы виды, потому что китайские палеонтологи считали их разными.[2]
История
Открытия
Макроолитус яйца были впервые обнаружены в Южном Китае китайским палеонтологом-пионером. Ян Чжунцзянь. Он описал останки нескольких окаменелых яиц из этого региона в 1965 году. Работая до появления современной паратаксономии окаменелых яиц, он дал им названия как виды Оолиты,[3] ныне несуществующее название, которое раньше использовалось для различных типов окаменелых яиц.[28][29] В 1975 году китайский палеонтолог Чжао Цзыкуй создал прототип современной паратаксономической системы, создав иерархическую систему семей, оогенов и видов. Чжао поместил Яна О. rugustus в новый ооген, Макроолитус, разделив его на два вида, М. yaotunensis и М. rugustus. Он также предположил, что американские виды "Оолит" карлиленский (который теперь классифицируется как другой ооген элонгатоолитид, Macroelongatoolithus[1]) рекомбинировать как Macroolithus carlylensis.[5]
В 1991 году русский палеонтолог Константин Михайлов представил современную классификацию окаменелых яиц, основанную на паратаксономической системе именования Чжао. Он классифицировал "О." Carlylensis в семье Spheroolithidae, но в остальном следовал классификации Чжао 1975 г. Макроолитус.[30] В 1994 году он назвал М. mutabilis, новый вид Макроолитус, основанный на останках, обнаруженных в Монголии.[2]
В 2000 г. появился четвертый вид, «М.» turolensis, был описан испанскими палеонтологами Ольгой Амо-Санхуан, Хосе Игнасио Канудо и Глорией Куэнка-Бескос на материалах из Испании. Однако, когда в 2014 г. был обнаружен новый материал этого вида, он был перемещен в собственный ооген, Guegoolithus, который в дальнейшем был перемещен в Spheroolithidae.[27]
В 2005 году яйца, возможно, относящиеся к М. yaotunensis были обнаружены в паре внутри таза ископаемого скелета овирапторид.[15] В 2008 году китайские палеонтологи Ченг Янь-нянь, Цзи Цян, У Сяо-чун и Шань Си-инь обнаружили пару яиц, которые являются первыми в Китае яйцами, содержащими эмбриональные останки овирапторозавров. Оба яйца были отнесены к М. yaotunensis.[19] Четвертый вид, М. lashuyuanensis, был описан в 2009 году китайскими палеонтологами Фанг Сяо-си, Ли Пэй-сянь, Чжан Чжи-цзюнь, Чжан Сянь-цю, Лин Ю-ли, Го Шэн-бин, Чэн Е-мин, Ли Чжэнь-ю, Чжан Сяо-цзюнь и Чэн Чжэн-у.[9]
Палеобиологические интерпретации

В 1994 году Михайлов предложил Макроолитус представляли собой яйца большого теропод, а именно Тарбозавр, исходя из большого размера яиц и в значительной степени перекрывающегося распределения.[2] Однако открытие эмбриона овирапторида внутри яйца элонгатоолитид поставило под сомнение эту гипотезу.[31] С открытием множественных ассоциаций овирапторозавров и элонгатоолитидов в конце 1990-х гг.[32] Яйца элонгатоолитов стали считаться принадлежащими динозаврам-овирапторозаврам.[1] Подтверждено происхождение овирапторидов Макроолитус когда в 2008 г. эмбрионы овирапторид, напоминающие Heyuannia были обнаружены внутри пары М. yaotunensis яйца в Цзянси.[19]
Вначале палеонтологи считали Макроолитус гнезда были закопаны. Mou 1992 отметил высокие значения газопроводности в Макроолитус яйца и поэтому пришел к выводу, что они были отложены в очень влажной среде, закопаны под землей или внутри насыпи.[33] Диминг (2006) нашел аналогичный результат. Это, кажется, противоречит свидетельствам того, что овирапторозавриды высиживали свои яйца, как птицы, но Диминг предположил, что Овираптор закопал яйца в кургане, а затем Овираптор и его родственники сидели на насыпи гнезда для инкубации, а не напрямую контактировали с яйцами.[34] Однако Танака и другие. (2015) критиковали эти результаты за отсутствие статистической строгости. На основе сравнения с современными яйцами они обнаружили, что Макроолитус прогнозировалось, что они откладываются в открытых или частично закрытых гнездах.[18] Wiemann и другие. (2017) также раскритиковали Му и Диминга за то, что они измерили пористость яичной скорлупы только в средней части яиц и не приняли во внимание тот факт, что плотность пор намного ниже вблизи полюсов. Это привело бы к завышению общей пористости яичной скорлупы и, следовательно, к завышению значения проводимости газа.[10]
Вымирание Macroolithus и других яиц из Южного Китая также имело историю различных интерпретаций. В 1990-х годах китайские палеонтологи, в том числе известный специалист по яйцам Чжао Цзыкуй, наблюдали постепенное сокращение разнообразия яиц динозавров в течение последних 200 000–300 000 лет мелового периода, при этом лишь Макроолитус вплоть до границы.[20][35] Они постулировали, вопреки гипотезе воздействия, что вымирание было результатом продолжительной засухи, которая увеличила концентрацию следов тяжелых металлов, что отрицательно сказалось на развитии яичной скорлупы и эмбрионов динозавров, что привело к постепенному сокращению и гибели популяции.[35] Чжао и другие. пересмотрел эту гипотезу в 2002 г., постулировав постепенное исчезновение Макроолитус вызвано вулканизмом Деканских ловушек.[21] В 2004 году Бак и другие. оспаривали эту интерпретацию, утверждая, что кажущееся постепенное вымирание было иллюзией, вызванной переработкой отложений.[24] Чжао и другие. (2009) утверждали, что вымирание было постепенным.[22]
Распространение и палеоэкология
Макроолитус известен из бесчисленных мест позднего мела в Китае, Монголии и Казахстане.[1]
Хэнань
В Хэнань, Macroolithus yaotunensis сосуществует с другими элонгатоолитами Elongatoolithus andrewsi и E. elongatus, а также Овалоолитус и Парасферолитус из Hugang, то Luyemiao, а Формации Сигу. Эти образования сформировались в позднем меловом периоде озерный или же болотный среда. Ископаемые останки динозавров редко встречаются в одних и тех же единицах, но троодонтиды, тираннозавры и гадрозавры известны из той же местности.[8]
Шаньдун
Некоторые яйца предварительно отнесены к Макроолитус известны из Группа Ванши в Лайян, Шаньдун.[36] Образования Группа Ванши были депонированы в аллювиальные вееры, плетеные каналы и мелкие озера. Общие окаменелости включают гадрозавридов и яйца динозавров.[37]
Южный Китай
М. rugustus, М. yaotunensis, и М. lashuyuanensis все известны из Бассейн Наньсюн в Гуандун.[5][9] Это образование было отложено в основном из ручьев, рек и озер в этом регионе.[21][35] Он охватывает K / T граница, после чего большая часть яиц динозавров исчезает, но Макроолитус очевидно, существует по обе стороны от границы (что означает, что некоторые виды динозавров дожили до раннего третичного периода).[21][22] Однако седиментологические данные свидетельствуют о том, что эти окаменелости были фактически переработаны потоками обломков в третичные породы.[24]
Бассейн Наньсюн известен обилием окаменелых яиц, преимущественно овирапторизавров.[1] яйца Elongatoolithus и Макроолитус.[21] Другие типы яиц включают других элонгатоолитидов, а также призматоолитиды, мегалоолиты, и овалоолитиды.[22] Следы показывают, что бассейн Наньсюн был населен орнитоподы, тероподы, и возможно зауроподы.[38]
Южный Китай, особенно Формация Наньсюн в Провинция Цзянси, возможно, имеет самое большое разнообразие овирапторозавров в мире,[39] а также включает несколько ассоциаций овирапторозавров с Макроолитус или аналогичные яйца.[19][15][14] Титанозавры и тираннозавриды также известны из этой области.[14] Фауна нединозавров включает ящериц и наземных nanhsiungchelyid черепахи.[40]
Монголия
Macroolithus rugustus в изобилии в Формация Немегт, который датируется поздним Кампанский рано Маастрихтский.[41][42] Это образование представляет собой отложения извилистой реки.[42] Хорошо сохранившиеся останки динозавров обычны в формации Немегт,[43] в том числе овирапторозавров,[42] титанозавры,[44] троодонтиды, тираннозавры, анкилозавры,[45] пахицефалозавры,[46] гадрозавры[47] орнитомимозавры, альваресзавры,[48] и теризинозавры.[49] Остатки мелких животных относительно редки, но несколько видов птиц известны из Немегта,[48] а также несколько видов многотуберкулезный млекопитающие.[50] Другие ископаемые яйца из формации Немегт включают Овалоолитус, Сфероолит, Elongatoolithus, и Laevisoolithus.[41]
М. mutabilis известен только в местности Их-Шунхт с Формация Барун Гойот, начиная с Сантон к Кампанский.[41][2] М. rugustus также известен из формации Барун-Гойот, которая представляет собой песчаные дюны, заполненные эоловый среда. По сравнению с формацией Немегт, крупные динозавры редки в Барун-Гойоте, где в фауне преобладают протоцератопсиды, овирапториды и анкилозавриды.[51] Другие типы окаменелых яиц из формации Барун-Гойот включают: Protoceratopsidovum, Gobioolithus, Фавеолоолитус, Дендроолит, Сфероолит, и Subtiliolithus.[41]
Казахстан
М. rugustus также был найден в Формация Манрак (также называемая Манракская Свита) Зайсанский бассейн в Восточно-Казахстанская область. Это образование находится недалеко от Река Тайжузген, и датируется некоторым временем в конце мелового периода, вероятно, маастриком.[2][4][31][52]
Смотрите также
Рекомендации
- ^ а б c d е ж грамм час я j k л м Саймон, Д. Дж. (2014). "Яйца гигантских динозавров (теропод) Oogenus Macroelongatoolithus (Elongatoolithidae) из юго-восточного Айдахо: таксономические, палеобиогеографические и репродуктивные последствия. "(Докторская диссертация, Государственный университет Монтаны, Бозман).
- ^ а б c d е ж грамм час я j k л Михайлов, Константин К. (1994). «Яйца теропод и протоцератопсов динозавров из мелового периода Монголии и Казахстана». Палеонтологический журнал. 28 (2): 101–120.
- ^ а б c d е Янг, Чжун-цзянь (1965). «Ископаемые яйца из Наньсюна, Гуантун и Канчжоу, Цзянси» (PDF). Позвоночные PalAsiatica. 9 (2): 141–170.
- ^ а б Михайлов, Константин (2000). «Яйца и скорлупа динозавров и птиц мелового периода Монголии». В Benton, M.J .; Шишкин, М.А .; Unwin, D.M .; Курочкин, Е. (ред.). Эпоха динозавров в России и Монголии. Издательство Кембриджского университета. С. 560–572.
- ^ а б c d е ж грамм час я Чжао, Цзыкуй (1975). «Микроструктура яичной скорлупы динозавров в Наньсюн, провинция Гуандун - по классификации яиц динозавров» (PDF). Позвоночные PalAsiatica. 13 (2): 105–117.
- ^ Карпентер, К. (1999). Яйца, гнезда и детеныши динозавров: взгляд на воспроизводство динозавров. Издательство Индианского университета. стр.294–295.
- ^ Карпентер, Кеннет (1999). «Как изучить ископаемое яйцо». Яйца, гнезда и детеныши динозавров: взгляд на воспроизводство динозавров (жизнь прошлого). Блумингтон, Индиана: Издательство Индианского университета. стр.122–144. ISBN 978-0-253-33497-8.
- ^ а б Лян, Синьцюань; Ван, Шуню; Ян, Дуншэн; Чжоу, Шицюань; Ву, Шичонг (2009). «Яйца динозавров и залежи яиц динозавров (верхний мел) в провинции Хэнань, Китай: проявления, палеоокружение, тафономия и сохранение». Прогресс естествознания. 19 (11): 1587–1601. Дои:10.1016 / j.pnsc.2009.06.012.
- ^ а б c d Фанг, Сяоси; Ли, Пэйсянь; Чжан, Чжицзюнь; Лин, Юли; Го, Шэнбинь; Ченг, Йеминг; Ли, Чжэньюй; Чжан, Сяоцзюнь; Ченг, Чжэнву (2009). "广东 南雄 白垩 系 及 恐龙 蛋 到 鸟蛋 演化 研究" [Меловые пласты в бассейне Наньсюн провинции Гуандун и эволюция от яйца динозавра до яйца птицы]. Acta Geoscientia Sinica (на китайском языке). 30 (2): 167–186.
- ^ а б c d Виманн, Жасмина; Ян, Цзы-Руэй; Sander, Philipp N .; Шнайдер, Марион; Энгезер, Марианна; Кат-Шорр, Стефани; Мюллер, Криста Э .; Сандер, Мартин П. (2017). «Динозавровое происхождение окраски яиц: овирапторы отложили сине-зеленые яйца». PeerJ. 5 (e3706): e3706. Дои:10.7717 / peerj.3706. ЧВК 5580385. PMID 28875070.
- ^ Банай, Г.Л. (1948). «Введение в медицинскую терминологию I. Греческие и латинские переводы». Бюллетень медицинской библиотечной ассоциации. 36 (1): 763–769. ЧВК 194697. PMID 16016791.
- ^ Михайлов, Константин Э .; Брей, Эмили С .; Хирш, Карл Ф. (1996). «Паратаксономия останков ископаемых яиц (Veterovata): принципы и приложения». Журнал палеонтологии позвоночных. 16 (4): 763–769. Дои:10.1080/02724634.1996.10011364.
- ^ Уайт, Джон Тахурдин (1858). Латинские суффиксы. Лондон: Spottiswoode and Co., стр.80.
латинский корень ensis.
- ^ а б c d е ж грамм Ван, Шо; Чжан, Шуканг; Салливан, Корвин; Сюй, Син (2016). «Яйца элонгатоолитов, содержащие эмбрионы овирапторид (Theropoda, Oviraptorosauria) из верхнего мела Южного Китая». BMC Эволюционная биология. 16 (67): 67. Дои:10.1186 / s12862-016-0633-0. ЧВК 4807547. PMID 27012653.
- ^ а б c d Сато, Тамаки; Чэн, Йен-нянь; У, Сяо-чун; Зеленицкий, Дарла; Сяо, Ю-фу (2005). "Пара яиц в скорлупе внутри самки динозавра". Наука. 308 (5720): 375. Дои:10.1126 / наука.1110578. PMID 15831749.
- ^ Fanti, F; Карри, П.Дж.; Бадамгарав, Д. (2012). «Новые образцы Немегтомайя из свит Баруунгойот и Немегт (поздний мел) Монголии ». PLoS ONE. 7 (2): e31330. Bibcode:2012PLoSO ... 731330F. Дои:10.1371 / journal.pone.0031330. ЧВК 3275628. PMID 22347465.
- ^ а б Варриккио, Дэвид Дж .; Мур, Джейсон Р .; Эриксон, Грегори М .; Норелл, Марк А .; Джексон, Фрэнки Д.; Борковский, Джон Дж. (2008). "Птичий отцовский уход произошел от динозавров". Наука. 322 (5909): 1826–1827. Bibcode:2008Научный ... 322.1826V. Дои:10.1126 / science.1163245. PMID 19095938.
- ^ а б Танака, Кохей; Зеленицкий, Дарла К .; Терриен, Франсуа (2015). «Пористость яичной скорлупы дает представление об эволюции гнездования динозавров». PLoS ONE. 10 (11): e0142829. Bibcode:2015PLoSO..1042829T. Дои:10.1371 / journal.pone.0142829. ЧВК 4659668. PMID 26605799.
- ^ а б c d е Чэн, Йен-нянь; Цзи, Цян; У, Сяо-чун; Шан, Си-у (2008). «Яйца овирапторозавров (динозавров) с эмбриональными скелетами, обнаруженные впервые в Китае». Acta Geologica Sinica. 82 (6): 1089–1094. Дои:10.1111 / j.1755-6724.2008.tb00708.x.
- ^ а б c d Чжао, Цзы-куй (1994). «Яйца динозавров в Китае: о структуре и эволюции яичной скорлупы». Карпентер, Кеннет; Хирш, Карл Ф .; Хорнер, Джон Р. (ред.). Яйца динозавров и младенцы. Издательство Кембриджского университета. С. 184–203. ISBN 978-0-521-44342-5.
- ^ а б c d е Чжао, Цзыкуй; Мао, Сюэминь; Чай, Чжифан; Ян, Гаочуан; Конг, Пинг; Эбихара, Мицуру; Чжао, Чжэньхуа (2002). «Возможная причинно-следственная связь между вымиранием динозавров и обогащением K / T иридия в бассейне Nanxiong, Южный Китай: свидетельства из яичной скорлупы динозавров». Палеогеография, палеоклиматология, палеоэкология. 178 (1–2): 1–17. Bibcode:2002ППП ... 178 .... 1Z. Дои:10.1016 / S0031-0182 (01) 00361-3.
- ^ а б c d Чжао, Цзыкуй; Мао, Сюэин; Чай, Чжифан; Ян, Гаочуан; Чжан, Фучэн; Ян, Чжэн (2009). «Геохимические изменения окружающей среды и вымирание динозавров во время перехода от мела к палеогену (K / T) в бассейне Наньсюн, Южный Китай: данные по скорлупе динозавров». Китайский научный бюллетень. 54 (5): 806. Дои:10.1007 / s11434-008-0565-1.
- ^ Ван, Цян; Ван, Сяолинь; Чжао, Цзыкуй (2010). «Недавний прогресс в изучении яиц динозавров в Китае» (PDF). Вестник Китайской академии наук. 24 (2): 95–97. Архивировано из оригинал (PDF) на 2017-01-09. Получено 2017-01-08.
- ^ а б c Бак, Бренда Дж .; Хэнсон, Эндрю Д .; Hengst, Richard A .; Ху, Шу-шэн (2004). ""Третичные динозавры "в бассейне Наньсюн, Южный Китай, переработаны из мелового периода". Журнал геологии. 112 (1): 111–118. Bibcode:2004JG .... 112..111B. Дои:10.1086/379695.
- ^ Varricctio, Дэвид Дж .; Барта, Дэниел Э. (2015). "Обновление" больших птичьих яиц "Сабата из мелового периода Гоби". Acta Palaeontologica Polonica. 60 (1): 11–25. Дои:10.4202 / app.00085.2014.
- ^ Гарсия, Джеральдин; Мариво, Лоран; Пелиси, Тьерри; Виани-Лиод, Моник (2006). «Ранняя яичная скорлупа зауроподов Лавриды» (PDF). Acta Palaeontologica Polonica. 51 (1): 99–104.
- ^ а б Морено-Азанза, М .; Canudo, J.I .; Гаска, Дж. М. (2014). «Яичная скорлупа сфероолитидов в нижнем меловом периоде Европы. Влияние на эволюцию яичной скорлупы у орнитисхийских динозавров» (PDF). Меловые исследования. 51: 75–87. Дои:10.1016 / j.cretres.2014.05.017.
- ^ Lawver, Daniel R .; Джексон, Фрэнки Д. (2015). "Обзор летописи окаменелостей размножения черепах: яйца, эмбрионы, гнезда и совокупляющиеся пары" (PDF). Бюллетень музея естественной истории Пибоди. 55 (2): 215–236. Дои:10.3374/014.055.0210.
- ^ Виани-Ляуд, Моник; Зеленицкий, Дарла (2003). «Исторические и новые взгляды на паратаксономию ископаемых яиц». Палеопозвоночные. 32 (2–4): 189–195.
- ^ Михайлов, Константин Э. (1991). "Классификация ископаемых яичных скорлуп амниотических позвоночных" (PDF). Acta Palaeontologica Polonica. 36 (2): 193–238.
- ^ а б Михайлов, К. (1997). «Ископаемая и современная яичная скорлупа амниотических позвоночных: тонкая структура, сравнительная морфология и классификация». Специальные статьи по палеонтологии. 56: 64.
- ^ Зеленицкий, Дарла К .; Карпентер, Кеннет; Карри, Филип Дж. (2000). "Первое обнаружение яичной скорлупы Elongatoolithus Theropod в Северной Америке: азиатский ооген Macroelongatoolithus из нижнего мела штата Юта ». Журнал палеонтологии позвоночных. 20 (1): 130–138. Дои:10.1671 / 0272-4634 (2000) 020 [0130: FROETE] 2.0.CO; 2.
- ^ Моу, Юн (1992). «Среда гнездования яиц позднемеловых динозавров из бассейна Наньсюн, провинция Гуандун» (PDF). Позвоночные PalAsiatica. 30 (2): 120–134.
- ^ Диминг, округ Колумбия (2006). «Ультраструктурная и функциональная морфология яичной скорлупы поддерживает идею, что яйца динозавров были инкубированы в субстрате». Палеонтология. 49 (1): 171–185. Дои:10.1111 / j.1475-4983.2005.00536.x.
- ^ а б c Чжао, Цзыкуй; Йе, Джи; Ли, Хуамэй; Чжао, Чжэньхуа; Ян, Чжэн (1991). «Вымирание динозавров через границу мелового и третичного периода в бассейне Наньсюн, провинция Гуандун» (PDF). Acta Paleontologica Sinica. 29 (1): 1–20.
- ^ Чжао, Цзыкуй (1979). «Прогресс в исследовании яиц динозавров» (PDF). Мезозойские и кайнозойские красные слои Южного Китая. Избранные статьи из «Семинара по меловому и третичному периоду», Наньсюн, провинция Гуандун: 330–340.
- ^ Ван, Цян; Ван, Сяолинь; Чжао, Цзыкуй; Чжан, Цзялян; Цзян, Шуньсин (2013). «Новая окаменелость яйца черепахи из верхнего мела бассейна Лайян, провинция Шаньдун, Китай». Anais da Academia Brasileira de Ciências. 85 (1): 103–111. Дои:10.1590 / S0001-37652013000100008. PMID 23538955.
- ^ Син, Лида; Харрис, Джеральд Д; Дун, Чжимин; Лин, Юли; Чен, Вэй; Го, Шэнбинь; Цзи, Цян (2009). «Следы орнитоподов (Dinosauria: Ornithischia) из формации Чжутянь верхнего мела в бассейне Наньсюн, провинция Гуандун, Китай, и общие наблюдения за большими китайскими следами орнитоподов» (PDF). Геологический бюллетень Китая. 28 (7): 829–843.
- ^ Люй Цзюньчан; Пу, Ханьюн; Кобаяси, Ёсицугу; Сюй, Ли; Чанг, Хуали; Шан, Юхуа; Лю, Ди; Ли, Юонг-Нам; Кундрат, Мартин; Шен, Caizhi (2015). «Новый динозавр овирапторид (Dinosauria: Oviraptorosauria) из позднего мела Южного Китая и его палеобиогеографические последствия». Научные отчеты. 5: 11490. Bibcode:2015НатСР ... 511490Л. Дои:10.1038 / srep11490. ЧВК 4489096. PMID 26133245.
- ^ Тонг, Хайянь; Мо, Джинью (2010). "Jiangxichelys, новая черепаха nanhsiungchelyid из позднего мела Ганьчжоу, провинция Цзянси, Китай". Геологический журнал. 147 (6): 981–986. Bibcode:2010ГеоМ..147..981Т. Дои:10.1017 / S0016756810000671.
- ^ а б c d Михайлов, Константин (1995). «Систематическое, фаунистическое и стратиграфическое разнообразие меловых яиц в Монголии: сравнение с Китаем». Шестой симпозиум по мезозойским наземным экосистемам и биоте, краткие доклады: 165–168.
- ^ а б c Weishampel, David B .; Фастовский, Давид Э .; Ватабе, Махито; Варриккио, Давид; Джексон, Фрэнки; Цогтбаатар, Хишигжав; Барсболд, Ринчен (2008). «Новые эмбрионы овирапторид из Бугин-Цав, свита Немегт (верхний мел), Монголия, с информацией об их среде обитания и росте». Журнал палеонтологии позвоночных. 28 (4): 1110–1119. Дои:10.1671/0272-4634-28.4.1110.
- ^ Кларк, Джулия А .; Норелл, Марк А. (2004). «Новые останки Авиалана и обзор известной орнитофауны из позднемеловой формации Немегт в Монголии». Американский музей Novitates. 3447: 1–12. Дои:10.1206 / 0003-0082 (2004) 447 <0001: NARAAR> 2.0.CO; 2. HDL:2246/2812.
- ^ Уилсон, Джеффри А. (2005). "Переописание монгольского зауропода" Nemegrosaurus mongoliensis Новинский (Dinosauria: Saurischia) и комментирует разнообразие зауроподов в позднем меловом периоде ». Журнал систематической палеонтологии. 3 (3): 283–318. Дои:10.1017 / S1477201905001628.
- ^ Беседка, Виктория М .; Карри, Филип Дж .; Бадамгарав, Демчиг (2014). «Анкилозавридные динозавры верхнемеловых отложений Баруунгойот и Немегт Монголии». Зоологический журнал Линнеевского общества. 172 (3): 631–652. Дои:10.1111 / zoj.12185.
- ^ Эванс, Дэвид С .; Браун, Калед Маршалл; Райан, Майкл Дж .; Цогтбаатар, Хишигжав (2011). "Черепная скульптура и онтогенетический статус Homalocephale calathocercos (Ornithischia: Pachycephalosauria) из формации Немегт, Монголия". Журнал палеонтологии позвоночных. 31 (1): 84–92. Дои:10.1080/02724634.2011.546287.
- ^ Марьянская, Тереза; Осмольска, Гальска (1982). «Фертский динозавр-ламбеозавр из формации Немегт, верхний мел, Монголия». Acta Palaeontologica Polonica. 26 (3/4): 243–255.
- ^ а б Кларк, Джулия А .; Норрелл, Марк А. (2004). «Новые останки Авиалана и обзор известной орнитофауны из позднемеловой формации Немегт в Монголии» (PDF). Американский музей Novitates. 3447: 1–12. Дои:10.1206 / 0003-0082 (2004) 447 <0001: NARAAR> 2.0.CO; 2. HDL:2246/2812.
- ^ Белл, Фил Р .; Карри, Филип Дж .; Ли, Юонг-Нам (2012). "Тиранозавр питается следами Дейнохейрус (Теропод:? Орнитомимозаврия) останки формации Немегт (поздний мел), Монголия ". Меловые исследования. 37: 186–190. Дои:10.1016 / j.cretres.2012.03.018.
- ^ Wible, John R .; Ружье, Гильермо В. (2000). "Черепная анатомия Криптобатор дашзевеги (Mammalia, Multituberculara) и его влияние на эволюцию характеров млекопитающих " (PDF). Бюллетень Американского музея естественной истории. 247: 6.
- ^ Лонгрич, Николас Р .; Карри, Филип Дж .; Донг, Чжи-Мин (2010). «Новый овирапторид (Dinosauria: Theropoda) из верхнего мела Баян Мандау, авторы Внутренней Монголии». Палеонтология. 53 (5): 945–960. Дои:10.1111 / j.1475-4983.2010.00968.x.
- ^ Аверьянов, Александр Олегович (2007). «Тероподные динозавры из позднекретакских отложений на северо-востоке Приаралья, Казахстан». Меловые исследования. 28 (3): 532–544. Дои:10.1016 / j.cretres.2006.08.008.