Ганглий дорсального корня - Dorsal root ganglion
| Ганглий дорсального корня | |
|---|---|
 Ганглии спинного корня (DRG) курицы эмбрион (примерно на стадии 7 дня) после инкубации в течение ночи в Среда роста NGF окрашены анти-нейрофиламент антитело. Видны невриты, растущие из ганглия. | |
 Спинной нерв с его вентральным и дорсальным корешками. Ганглии дорзального корешка - это «спинномозговой ганглий», следующий за спинным корешком. | |
| Подробности | |
| Предшественник | нервный гребень |
| Идентификаторы | |
| латинский | ганглии сенсориума нервов спинного мозга |
| MeSH | D005727 |
| TA98 | A14.2.00.006 |
| TA2 | 6167 |
| FMA | 5888 |
| Анатомическая терминология | |
А ганглий дорсального корня (или же спинномозговой узел; также известный как задний корневой ганглий) представляет собой кластер нейроны (а ганглий ) в дорсальный корешок из спинной нерв. Клеточные тела сенсорных нейронов, известные как нейроны первого порядка расположены в ганглиях задних корешков.[1]
В аксоны нейронов ганглия дорсального корешка известны как афференты. в периферическая нервная система, афференты относятся к аксонам, которые передают сенсорную информацию в Центральная нервная система (т.е. мозг и спинной мозг ).
Структура
Нейроны, составляющие ганглий дорзального корешка, относятся к псевдо-униполярный типа, то есть у них есть тело клетки (сома) с двумя ветвями, которые действуют как один аксон, часто называемый дистальный отросток и проксимальный отросток.
В отличие от большинства нейронов, обнаруженных в Центральная нервная система, потенциал действия нейрон заднего корневого ганглия может инициироваться в дистальный отросток на периферии, обходят тело клетки и продолжают распространяться по проксимальный отросток до достижения синаптический терминал в задний рог спинного мозга.
Дистальный разрез
Дистальный участок аксона может быть либо оголенным нервным окончанием, либо инкапсулирован структурой, которая помогает передавать конкретную информацию нерву. Два примера, когда нервное окончание дистальный отросток инкапсулирован как таковой, Тельца Мейснера, которые отображают дистальные отростки механосенсорных нейронов, чувствительных только к поглаживанию, и Тельца Пачини, которые делают нейроны более чувствительными к вибрации.[2]
Место расположения
Ганглии дорзальных корешков лежат в межпозвонковые отверстия. Передний и задний корешки спинномозговых нервов соединяются сразу за (латеральным) местом расположения ганглия задних корешков.
Разработка
Ганглии дорзальных корешков развиваются у эмбриона из нервный гребень клетки, а не нервная трубка. Следовательно, спинномозговые ганглии можно рассматривать как серое вещество спинного мозга, переместившееся на периферию.
Функция
Ноцицепция
Протон-чувствительные рецепторы G-белка экспрессируются сенсорными нейронами DRG и могут играть роль в кислотно-индуцированном ноцицепция.[3]
Механочувствительные каналы
Нервные окончания нейронов ганглия задних корешков имеют множество сенсорных рецепторов, которые активируются механическими, термическими, химическими и ядовитыми раздражителями.[4] В этих сенсорных нейронах группа ионные каналы считается ответственным за соматосенсорный трансдукции. Сжатие ганглия дорзального корешка механическим раздражителем снижает порог напряжения, необходимый для вызова реакции, и вызывает потенциалы действия быть уволенным.[5] Это возбуждение может сохраняться даже после устранения раздражителя.[5]
Два разных типа механочувствительные ионные каналы были обнаружены в нейронах ганглия заднего корешка. Эти два канала в целом классифицируются как с высоким порогом (HT) или как с низким порогом (LT).[4] Как следует из их названий, у них разные пороги, а также разная чувствительность к давлению. Это катионные каналы, активность которых, по-видимому, регулируется правильным функционированием цитоскелета и белков, связанных с цитоскелетом.[4] Наличие этих каналов в ганглии заднего корешка дает основание полагать, что другие сенсорные нейроны также могут содержать их.
Высокопороговые механочувствительные каналы
Каналы с высоким порогом могут сыграть важную роль в ноцицепция. Эти каналы обнаруживаются преимущественно в более мелких сенсорных нейронах ганглиозных клеток дорсального корешка и активируются более высоким давлением - два признака, характерных для ноцицепторов.[4] Кроме того, порог HT-каналов был снижен в присутствии PGE2 (соединение, которое сенсибилизирует нейроны к механическим стимулам и механической гипералгезии), что дополнительно подтверждает роль HT-каналов в преобразовании механических стимулов в ноцицептивные нейронные сигналы.[4][5][6]
Пресинаптический контроль
Пресинаптическая регуляция отделяемого окончания спинного нерва в спинном мозге может происходить посредством определенных типов ГАМКА рецепторы но не через активацию рецепторы глицина которые отсутствуют в терминалах этих типов. Таким образом ГАМКА рецепторы но нет рецепторы глицина может пресинаптически контролировать ноцицепция и боль коробка передач.[7]
Смотрите также
Рекомендации
- ^ Первес, Дейл; Августин, Джордж Дж .; Фитцпатрик, Дэвид; Кац, Лоуренс С .; Ламантия, Энтони-Самуэль; Макнамара, Джеймс О .; Уильямс, С. Марк (2001). «Главный афферентный путь для механосенсорной информации: система дорсального столба-медиального лемниска». Неврология. 2-е издание. Получено 30 мая 2018.
- ^ Кандел ER, Шварц JH, Джесселл TM. Принципы нейронологии, 4-е изд., С.431–433. Макгроу-Хилл, Нью-Йорк (2000). ISBN 0-8385-7701-6
- ^ Хуанг CW, Цзэн Дж.Н., Чен Ю.Дж., Цай В.Ф., Чен С.К., Сун WH (2007). «Ноцицепторы ганглия задних корешков экспрессируют протон-чувствительные рецепторы, связанные с G-белком». Мол. Клетка. Неврологи. 36 (2): 195–210. Дои:10.1016 / j.mcn.2007.06.010. PMID 17720533.
- ^ а б c d е Чо, H .; Шин, Дж .; Shin, C. Y .; Lee, S. Y .; О, У. (2002). «Механочувствительные ионные каналы в культивируемых сенсорных нейронах новорожденных крыс». Журнал неврологии. 22 (4): 1238–1247. PMID 11850451.
- ^ а б c Sugawara, O .; Atsuta, Y .; Iwahara, T .; Мурамото, Т .; Watakabe, M .; Такемицу, Ю. (1996). «Влияние механического сжатия и гипоксии на нервный корешок и ганглии задних корешков. Анализ эктопического возбуждения с использованием модели in vitro». Позвоночник. 21 (18): 2089–2094. Дои:10.1097/00007632-199609150-00006. PMID 8893432.
- ^ Syriatowicz, J. P .; Hu, D .; Уокер, Дж. С .; Трейси, Д. Дж. (1999). «Гипералгезия из-за повреждения нерва: роль простагландинов». Неврология. 94 (2): 587–594. Дои:10.1016 / S0306-4522 (99) 00365-6. PMID 10579219.
- ^ Лоренцо Л.Е., Годин А.Г., Ван Ф., Сент-Луис М., Карбонетто С., Уайзман П.В., Рибейро-да-Силва А., Де Конинк Ю. (июнь 2014 г.). «Кластеры гефирина отсутствуют в первичных афферентных окончаниях малого диаметра, несмотря на присутствие ГАМК.А Рецепторы ». J. Neurosci. 34 (24): 8300–17. Дои:10.1523 / JNEUROSCI.0159-14.2014. PMID 24920633.
Дополнительные изображения
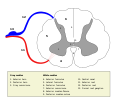
Спинной мозг
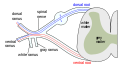
Формирование спинномозгового нерва из заднего и переднего корешков
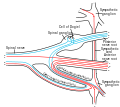
Схема, показывающая структуру типичного спинномозгового нерва.



внешняя ссылка
- Анатомическая фигура: 02: 04-09 в Human Anatomy Online, Медицинский центр SUNY Downstate
- Гистологическое изображение: 04401loa - Система обучения гистологии в Бостонском университете
- Фото модели в Государственный университет Огайо
- Диаграмма на webanatomy.net
- Фото на uwlax.edu
{kind=link}
| Авторитетный контроль |
|---|