Гель-электрофорез нуклеиновых кислот - Gel electrophoresis of nucleic acids


Электрофорез нуклеиновой кислоты аналитический метод, используемый для разделения ДНК или же РНК осколки по размеру и реакционной способности. Молекулы нуклеиновой кислоты, которые должны быть проанализированы, помещены в вязкую среду, гель, где электрическое поле индуцирует нуклеиновые кислоты (которые заряжены отрицательно из-за их сахаристости).фосфат позвоночника), чтобы перейти к анод (который заряжен положительно, потому что это электролитический скорее, чем гальванический элемент ). Разделение этих фрагментов достигается за счет использования подвижностей, с которыми молекулы разных размеров могут проходить через гель. Более длинные молекулы мигрируют медленнее, потому что они испытывают большее сопротивление внутри геля. Поскольку размер молекулы влияет на ее подвижность, более мелкие фрагменты оказываются ближе к аноду, чем более длинные за данный период. Через некоторое время напряжение снимают и анализируют градиент фрагментации. Для больших расстояний между фрагментами одинакового размера можно увеличить напряжение или время работы. Длительные исследования геля низкого напряжения обеспечивают наиболее точное разрешение. Однако напряжение не является единственным фактором при определении электрофореза нуклеиновых кислот.
Разделяемую нуклеиновую кислоту можно получить несколькими способами перед разделением с помощью электрофореза. В случае больших молекул ДНК, ДНК часто разрезают на более мелкие фрагменты с помощью ДНК. эндонуклеаза рестрикции (или рестрикционный фермент). В других случаях, например, ПЦР В амплифицированных образцах ферменты, присутствующие в образце, которые могут повлиять на разделение молекул, перед анализом удаляются различными способами. После того, как нуклеиновая кислота подготовлена должным образом, образцы раствора нуклеиновой кислоты помещают в лунки геля, и на гель прикладывают напряжение в течение определенного времени.
Фрагменты ДНК разной длины визуализируются с помощью флуоресцентный краситель специфичен для ДНК, например этидиум бромид. Гель показывает полосы, соответствующие различным популяциям молекул нуклеиновых кислот с разной молекулярной массой. Размер фрагмента обычно указывается в «нуклеотидах», «парах оснований» или «т.п.н.» (для тысяч пар оснований) в зависимости от того, была ли разделена одно- или двухцепочечная нуклеиновая кислота. Определение размера фрагмента обычно проводится путем сравнения с коммерчески доступными Маркеры ДНК содержащие линейные фрагменты ДНК известной длины.
Типы гелей, наиболее часто используемых для электрофореза нуклеиновых кислот: агароза (для относительно длинных молекул ДНК) и полиакриламид (для высокого разрешения коротких молекул ДНК, например в Секвенирование ДНК ). Гели обычно запускались в формате «плиты», таком как показано на рисунке, но капиллярный электрофорез стало важным для таких приложений, как высокопроизводительное секвенирование ДНК. Методы электрофореза, используемые при оценке Повреждение ДНК включают щелочной гель-электрофорез и гель-электрофорез в импульсном поле.
Для коротких сегментов ДНК, таких как двухцепочечная ДНК от 20 до 60 п.н., обработка их в полиакриламидном геле (PAGE) даст лучшее разрешение (нативное состояние).[1] Точно так же РНК и одноцепочечную ДНК можно запускать и визуализировать с помощью гелей для ПААГ, содержащих денатурирующие агенты, такие как мочевина. PAGE гели широко используются в таких техниках, как печать стопы ДНК, EMSA и другие методы взаимодействия ДНК-белок.
Измерение и анализ в основном выполняются с помощью специального программного обеспечения для анализа гелей. Результаты капиллярного электрофореза обычно отображаются в виде графика, который называется электрофореграмма.
Факторы, влияющие на миграцию нуклеиновых кислот
На миграцию нуклеиновых кислот может влиять ряд факторов: размер пор геля, используемое напряжение, ионная сила буфера и концентрация. вставка краситель, такой как бромистый этидий, если используется во время электрофореза.[2]
Размер ДНК
Гель просеивает ДНК размером с молекулу ДНК, благодаря чему более мелкие молекулы перемещаются быстрее. Двуцепочечная ДНК движется со скоростью, примерно обратно пропорциональной скорости движения. логарифм количества пар оснований. Однако это соотношение нарушается с очень большими фрагментами ДНК, и их невозможно разделить с помощью стандартных электрофорез в агарозном геле. Предел разрешения зависит от состава геля и напряженности поля.[3] и на подвижность более крупной кольцевой ДНК может сильнее влиять размер пор геля, чем на линейную ДНК.[4] Разделение очень больших фрагментов ДНК требует гель-электрофорез в импульсном поле (PFGE). В гель-электрофорезе с инверсией поля (FIGE, разновидность PFGE) возможна «инверсия полосы», когда большие молекулы могут двигаться быстрее, чем маленькие.
Конформация ДНК
В конформация молекулы ДНК может существенно повлиять на движение ДНК, например, сверхспиральная ДНК обычно движется быстрее, чем расслабленная ДНК, потому что она плотно свернута и, следовательно, более компактна. В нормальном препарате плазмидной ДНК может присутствовать несколько форм ДНК,[5] и гель, полученный при электрофорезе плазмид, обычно показывает основную полосу, которая является отрицательно свернутой формой, тогда как другие формы ДНК могут проявляться как второстепенные более слабые полосы. Эти второстепенные полосы могут быть ДНК с разрывами (открытая круговая форма) и расслабленной закрытой круговой формой, которые обычно работают медленнее, чем сверхспиральная ДНК, а одноцепочечная форма (которая иногда может появляться в зависимости от методов приготовления) может двигаться впереди суперспиральной ДНК. Однако скорость, с которой перемещаются различные формы, может изменяться при использовании различных условий электрофореза, например, линейная ДНК может работать быстрее или медленнее, чем суперспиральная ДНК, в зависимости от условий.[6] и на подвижность более крупной кольцевой ДНК может сильнее влиять размер пор геля, чем на линейную ДНК.[4] Если не суперспиральный Маркеры ДНК используются, размер кольцевой ДНК-подобной плазмиды может быть более точно определен после ее линеаризации с помощью ограничительный дайджест.
Повреждение ДНК из-за повышенного сшивание также будет уменьшать электрофоретическую миграцию ДНК в зависимости от дозы.[7][8]
Концентрация бромистого этидия
На кольцевую ДНК сильнее влияет концентрация бромида этидия, чем на линейную ДНК, если бромид этидия присутствует в геле во время электрофореза. Все естественные круги ДНК скручены, но бромид этидия, который внедряется в кольцевую ДНК, может изменять заряд, длину, а также суперсильность молекулы ДНК, поэтому его присутствие во время электрофореза может повлиять на ее движение в геле. Увеличение количества бромида этидия, внедренного в ДНК, может превратить ее из отрицательно свернутой в спираль молекулы в полностью расслабленную форму, а затем в положительно свернутую суперспираль при максимальной интеркаляции.[9] Электрофорез в агарозном геле можно использовать для разделения кольцевой ДНК с различной топологией суперспирализации.
Концентрация геля
Концентрация геля определяет размер пор геля, который влияет на миграцию ДНК. Разрешение ДНК изменяется в зависимости от процентной концентрации геля. Увеличение концентрации агарозы в геле снижает скорость миграции и улучшает разделение более мелких молекул ДНК, в то время как снижение концентрации геля позволяет разделять большие молекулы ДНК. Для стандартного электрофореза в агарозном геле 0,7% дает хорошее разделение или разрешение больших фрагментов ДНК размером 5–10 килобайт, а 2% гель дает хорошее разрешение для небольших фрагментов размером 0,2–1 килобайт. До 3% можно использовать для разделения очень мелких фрагментов, но вертикальный полиакриламидный гель больше подходит для разрешения мелких фрагментов. Однако гель с высокой концентрацией требует более длительного времени работы (иногда дней), а гели с высоким процентным содержанием часто бывают хрупкими и могут неравномерно затвердеть. Гели с высоким процентным содержанием агарозы следует запускать с PFGE или FIGE. Гели с низким процентным содержанием (0,1-0,2%) хрупкие и могут сломаться. 1% гели широко используются во многих областях.[10]
Прикладное поле
При низких напряжениях скорость миграции ДНК пропорциональна приложенному напряжению, то есть чем выше напряжение, тем быстрее движется ДНК. Однако с увеличением напряженности электрического поля подвижность высокомолекулярных фрагментов ДНК дифференциально возрастает, и эффективный диапазон разделения уменьшается, и поэтому разрешение уменьшается при высоком напряжении. Для оптимального разрешения ДНК размером более 2 КБ при стандартном гель-электрофорезе рекомендуется от 5 до 8 В / см.[6] Напряжение также ограничено тем фактом, что оно нагревает гель и может вызвать плавление геля, если гель работает под высоким напряжением в течение длительного периода, особенно для геля агарозы с низкой температурой плавления.
Однако подвижность ДНК может изменяться в неустойчивом поле. В поле, которое периодически меняют, подвижность ДНК определенного размера может значительно снизиться при определенной частоте циклов.[11] Это явление может привести к инверсии полосы, в результате чего более крупные фрагменты ДНК перемещаются быстрее, чем более мелкие в PFGE.
Механизм миграции и отделения
Отрицательный заряд его фосфатного скелета перемещает ДНК к положительно заряженному аноду во время электрофореза. Однако миграция молекул ДНК в растворе в отсутствие гелевой матрицы не зависит от молекулярной массы во время электрофореза, то есть без гелевой матрицы невозможно разделение по размеру.[12] Гидродинамическое взаимодействие между различными частями ДНК прерывается потоком противоионов, движущимся в противоположном направлении, поэтому не существует механизма для создания зависимости скорости от длины в масштабе, превышающем длину экранирования около 10 нм.[11] Это отличает его от других процессов, таких как седиментация или диффузия, где важно гидродинамическое взаимодействие на больших расстояниях.
Таким образом, гелевая матрица отвечает за разделение ДНК по размеру во время электрофореза, однако точный механизм, ответственный за разделение, не совсем ясен. Существует ряд моделей для механизма разделения биомолекул в гелевой матрице, широко распространенной является модель Огстона, которая рассматривает полимерную матрицу как сито, состоящее из случайно распределенной сети связанных между собой пор.[13] Глобулярный белок или случайный катушки ДНК движется через соединенные поры, достаточно большие, чтобы обеспечить ее прохождение, и движение более крупных молекул с большей вероятностью будет затруднено и замедлено из-за столкновений с гелевой матрицей, поэтому молекулы разных размеров могут быть разделены в этом процессе просеивания. .[11]
Однако модель Огстона не работает для больших молекул, в результате чего поры значительно меньше размера молекулы. Для молекул ДНК размером более 1 т.п.н. отказ модель (или ее варианты) наиболее часто используются. Эта модель предполагает, что ДНК может ползать «змееподобным» образом (отсюда и «рептация») через поры в виде удлиненной молекулы. При более высокой напряженности электрического поля это превратилось в смещенную модель отражения, в результате чего передний конец молекулы становится сильно смещенным в прямом направлении, и этот передний край тянет за собой остальную часть молекулы. В режиме полного смещения подвижность достигла точки насыщения, и ДНК сверх определенного размера не может быть отделена.[13] Однако идеальное параллельное выравнивание цепи с полем на практике не наблюдается, поскольку это означало бы одинаковую подвижность для длинных и коротких молекул.[11] Дальнейшее уточнение модели смещенной рептации учитывает внутренние колебания цепи.[14]
Смещенная модель рептации также использовалась для объяснения подвижности ДНК в PFGE. Ориентация ДНК постепенно создается рептацией после начала действия поля, и время, в течение которого она достигает стационарной скорости, зависит от размера молекулы. Когда поле изменяется, более крупным молекулам требуется больше времени для переориентации, поэтому можно отличить длинные цепи, которые не могут достичь своей установившейся скорости, от коротких, которые большую часть времени перемещаются с постоянной скоростью.[14] Однако существуют и другие модели.
Флуоресцентная микроскопия окрашенных молекул в реальном времени показала более тонкую динамику во время электрофореза, при этом ДНК демонстрирует значительную эластичность, поскольку она попеременно растягивается в направлении приложенного поля, а затем сжимается в шар или зацепляется в U-образную форму, когда она зацепились за полимерные волокна.[15][16] Это наблюдение можно назвать «гусеничной» моделью.[17] Другая модель предполагает, что ДНК сцепляется с полимерной матрицей, и чем больше молекула, тем больше вероятность, что она запутается и ее движение будет затруднено.[18]
Визуализация

Наиболее распространенный краситель, используемый для того, чтобы сделать полосы ДНК или РНК видимыми для электрофореза в агарозном геле, - это этидиум бромид, обычно сокращенно EtBr. Он флуоресцирует в ультрафиолетовом свете при внедрении в большую бороздку ДНК (или РНК). Пропуская ДНК через гель, обработанный EtBr, и визуализируя ее в УФ-свете, можно отчетливо видеть любую полосу, содержащую более ~ 20 нг ДНК. EtBr - известный мутаген,[19] и доступны более безопасные альтернативы, такие как GelRed, произведено Biotium, который прикрепляется к малой бороздке.[20]
SYBR Зеленый I это еще один краситель дцДНК, продуцируемый Invitrogen. Он дороже, но в 25 раз более чувствителен и, возможно, безопаснее, чем EtBr, хотя нет данных, касающихся его мутагенности или токсичности для людей.[21]
SYBR Сейф представляет собой вариант SYBR Green, обладающий достаточно низкими уровнями мутагенности и токсичности, чтобы считаться неопасными отходами в соответствии с федеральными правилами США.[22] Он имеет аналогичные уровни чувствительности к EtBr,[22] но, как и SYBR Green, значительно дороже. Однако в странах, где безопасное удаление опасных отходов является обязательным, затраты на утилизацию EtBr могут легко превысить первоначальную разницу в цене.
Поскольку окрашенная EtBr ДНК не видна в естественном свете, ученые смешивают ДНК с отрицательно заряженными загрузка буферов перед добавлением смеси в гель. Буферы загрузки полезны, потому что они видны в естественном свете (в отличие от УФ-света для ДНК, окрашенной EtBr), и они осаждаются совместно с ДНК (что означает, что они движутся с той же скоростью, что и ДНК определенной длины). Ксилол цианол и Бромфеноловый синий обычные красители, содержащиеся в загрузочных буферах; они работают примерно с той же скоростью, что и фрагменты ДНК длиной 5000 и 300 пар оснований соответственно, но их точное положение зависит от процентного содержания геля. Другие менее часто используемые маркеры прогресса: Крезол Красный и Оранжевый G которые составляют примерно 125 и 50 п.н. соответственно.
Визуализации также можно добиться путем переноса ДНК после SDS-СТРАНИЦА к нитроцеллюлозной мембране с последующим воздействием гибридизационный зонд. Этот процесс называется Саузерн-блоттинг.
Для флуоресцентных красителей после электрофореза гель освещают ультрафиолетовый лампа (обычно помещая ее на световой короб, используя защитное снаряжение для ограничения воздействия ультрафиолетового излучения). Осветительное устройство в основном также содержит устройство формирования изображения, которое делает изображение геля после освещения УФ-излучением. В этидиум бромид флуоресценция красновато-оранжевый в присутствии ДНК, так как она интеркалирована с ДНК. Полосу ДНК также можно вырезать из геля и затем растворить, чтобы извлечь очищенную ДНК. Затем гель можно сфотографировать, как правило, с помощью цифровой или поляроидной камеры. Хотя окрашенная нуклеиновая кислота имеет красновато-оранжевый цвет, изображения обычно отображаются в черно-белом цвете (см. Рисунки). Повреждение образца ДНК ультрафиолетом может снизить эффективность последующих манипуляций с образцом, таких как лигирование и клонирование. Ультрафиолетовое излучение с более короткой длиной волны (302 или 312 нм) вызывает больший ущерб, например, воздействие всего за 45 секунд может значительно уменьшить эффективность трансформации. Следовательно, если ДНК будет использоваться для последующих процедур, следует ограничить воздействие УФ-излучения с более короткой длиной волны, вместо этого следует использовать УФ-излучение с более высокой длиной волны (365 нм), которое причиняет меньший ущерб. Однако излучение с более высокой длиной волны вызывает более слабую флуоресценцию, поэтому, если необходимо захватить изображение геля, можно использовать более коротковолновый УФ-свет в течение короткого времени. Добавление Цитидин или же гуанозин в буфер для электрофореза в концентрации 1 мМ может защитить ДНК от повреждения.[23] В качестве альтернативы, источник возбуждения синего света с возбуждаемым синим пятном, например SYBR Зеленый или же GelGreen может быть использовано.
При исследовании гель-электрофореза часто используются программные инструменты анализа изображений, такие как ImageJ.
| 1 | 2 | 3 |
|---|---|---|
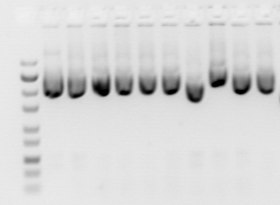
 1% агарозный «пластинчатый» гель при нормальном освещении за УФ-экраном из плексигласа. Видны только маркерные красители |  Гель с УФ-подсветкой, этидиум бромид окрашенная ДНК светится оранжевым |  Цифровое фото геля. Дорожка 1. Коммерческие ДНК-маркеры (1kbplus), дорожка 2. пустая, дорожка 3. а ПЦР продукт чуть более 500 оснований, дорожка 4. Ограничение дайджест, показывающий аналогичный фрагмент, вырезанный из 4,5 кб плазмида вектор |
Рекомендации
- ^ Джагува Васудеван, Ананда Айяппан; Марио Перкович; Янник Буллиард; Клаус Чичутек; Дидье Троно; Дитер Хойссингер; Карстен Мюнк (август 2013 г.). «Ставка на прототип пенистого вируса ухудшает димеризацию и цитозольную растворимость человеческого APOBEC3G». Журнал вирусологии. 87 (16): 9030–9040. Дои:10.1128 / JVI.03385-12. ЧВК 3754047. PMID 23760237.
- ^ Г. Люкотт; Ф. Банейкс (1993). Введение в методы молекулярного клонирования. Вили-Блэквелл. п. 32. ISBN 978-0471188490.
- ^ Джозеф Сэмбрук; Дэвид Рассел. «Глава 5, протокол 1». Молекулярное клонирование - лабораторное руководство. 1 (3-е изд.). п. 5.2. ISBN 978-0-87969-577-4.
- ^ а б Aaij C, Borst P (1972). «Гель-электрофорез ДНК». Biochim Biophys Acta. 269 (2): 192–200. Дои:10.1016/0005-2787(72)90426-1. PMID 5063906.
- ^ Ричард Р. Синден (24 ноября 1994 г.). Структура и функции ДНК. Academic Press Inc. стр. 97. ISBN 978-0126457506.
- ^ а б Джозеф Сэмбрук; Дэвид Рассел. «Глава 5, протокол 1». Молекулярное клонирование - лабораторное руководство. 1 (3-е изд.). С. 5.5–5.6. ISBN 978-0-87969-577-4.
- ^ Blasiak J, Trzeciak A, Malecka-Panas E, Drzewoski J, Wojewódzka M (2000). «Генотоксичность этанола и ацетальдегида in vitro в лимфоцитах человека и клетках слизистой оболочки желудочно-кишечного тракта». Токсикология in vitro. 14 (4): 287–295. Дои:10.1016 / S0887-2333 (00) 00022-9. PMID 10906435.
- ^ Лу И, Моримото К. (2009). «Связано ли обычное употребление алкоголя со снижением миграции электрофоретической ДНК в лейкоцитах периферической крови у мужчин японцев с дефицитом ALDH2?». Мутагенез. 24 (4): 303–308. Дои:10.1093 / mutage / gep008. PMID 19286920.
- ^ Дональд Воет; Джудит Г. Воет (1995). Биохимия (2-е изд.). Джон Вили и сыновья. стр.877–878. ISBN 978-0471586517.
- ^ «Электрофорез в агарозном геле (основной метод)». Биологические протоколы. Получено 23 августа 2011.
- ^ а б c d Зимм Б. Х., Левен С. Д. (1992). «Проблемы и перспективы теории гель-электрофореза ДНК» (PDF). Ежеквартальные обзоры биофизики. 25 (2): 171–204. Дои:10,1017 / с0033583500004662. PMID 1518924.
- ^ Роберт В. Олд; Сэнди Б. Примроуз (27 сентября 1994 г.). Принцип манипуляции генами - введение в генную инженерию (5-е изд.). Blackwell Scientific. п.9. ISBN 9780632037124.
- ^ а б Ли Чжу; Хун Ван (2 марта 2009 г.). «Глава 4 - Генетический анализ в миниатюрных системах электрофореза». Инь Тянь, Вэй-Чэн; Finehout, Эрин (ред.). Микрофлюидика для биологических применений. Springer. п. 125. ISBN 978-0-387-09480-9.
- ^ а б Жан-Луи Виови (2000). «Электрофорез ДНК и других полиэлектролитов: физические механизмы». Обзоры современной физики. 72 (3): 813–872. Bibcode:2000РвМП ... 72..813В. Дои:10.1103 / RevModPhys.72.813.
- ^ Смит С.Б., Олдридж П.К., Каллис Дж. Б. (1989). «Наблюдение за отдельными молекулами ДНК, подвергающимися гель-электрофорезу». Наука. 243 (4888): 203–206. Bibcode:1989Sci ... 243..203S. Дои:10.1126 / science.2911733. PMID 2911733.
- ^ Шварц Д.К., Коваль М (1989). «Конформационная динамика отдельных молекул ДНК при гель-электрофорезе». Природа. 338 (6215): 520–2. Bibcode:1989Натура.338..520С. Дои:10.1038 / 338520a0. PMID 2927511.
- ^ Дэвид Шихан (2009), Физическая биохимия: принципы и применение (2-е изд.), Wiley-Blackwell, стр. 181, ISBN 978-0470856031
- ^ Форстер Р.Э., Херт Д.Г., Кизл Т.Н., Фредлейк С.П., Баррон А.Е. (2009). «Анализ механизма миграции ДНК для применения в капиллярном и микрочиповом электрофорезе». Электрофорез. 30 (12): 2014–24. Дои:10.1002 / elps.200900264. ЧВК 2762034. PMID 19582705.
- ^ Бегусова, М; и другие. (2000). «Влияние интеркаляции бромистого этидия на радиочувствительность ДНК». Int J Radiat Biol. 76 (1). Явное использование et al. в:
| last1 =(помощь) - ^ [1]
- ^ «Гель-краситель на основе нуклеиновой кислоты SYBR Green I» (PDF). Архивировано из оригинал (PDF) на 2012-05-22. Получено 2013-06-23.
- ^ а б «Гель-краситель SYBR Safe DNA» (PDF). Архивировано из оригинал (PDF) на 2012-09-07. Получено 2013-06-23.
- ^ Грюндеманн Д., Шёмиг Э. (1996). «Защита ДНК во время препаративного электрофореза в агарозном геле от повреждений, вызванных ультрафиолетом» (PDF). Биотехнологии. 21 (5): 898–903. Дои:10.2144 / 96215rr02. PMID 8922632. Архивировано из оригинал (PDF) на 2016-03-04. Получено 2017-11-26.