Морской еж - Википедия - Sea urchin
| Морской еж | |
|---|---|
 | |
| Tripneustes ventricosus и Echinometra viridis | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Иглокожие |
| Подтип: | Echinozoa |
| Учебный класс: | Echinoidea Леске, 1778 |
| Подклассы | |
| |
Морские ежи (/ˈɜːrtʃɪпz/), обычно колючий, шаровидный животные, иглокожие в классе Echinoidea. Около 950 видов обитают на морском дне, населяя все океаны и глубинные зоны от приливной зоны до 5000 метров (16000 футов; 2700 саженей).[1] Их тесты (твердые раковины) круглые и колючие, обычно от 3 до 10 см (от 1 до 4 дюймов) в поперечнике. Морские ежи передвигаются медленно, ползая трубка ноги, а иногда и толкаются шипами. Питаются в основном водоросли но также ешьте медленные или сидячий животные. Их хищники включают морские выдры, морская звезда, волк угорь, спинорог, и люди.
Как и другие иглокожие, ежи имеют пятикратную симметрию, как взрослые особи, но их личинки плютеуса имеют двусторонняя (зеркальная) симметрия, указывая, что они принадлежат Bilateria, большая группа животных тип это включает хордовые, членистоногие, кольчатые червя и моллюски. Они широко распространены во всех океанах, во всех климатах от тропического до полярного, и населяют морские бентические (морское дно) местообитания от скалистых берегов до хадальная зона глубины. У ехиноидов есть богатая летопись окаменелостей, относящаяся к Ордовик около 450 миллионов лет назад. Их ближайшими родственниками среди иглокожих являются морские огурцы (Holothuroidea); оба дейтеростомы, клада, которая включает хордовые.
Животные изучаются с 19 века как модельные организмы в биология развития, поскольку их эмбрионы было легко наблюдать; это продолжалось исследованиями их геномы из-за их необычной пятикратной симметрии и отношения к хордовым. Такие виды, как грифельный карандаш еж популярны в аквариумах, где они полезны для борьбы с водорослями. Ископаемые ежи использовались в качестве защитных амулеты.
Разнообразие
Морские ежи являются членами филюм Иглокожие, в который также входят морские звезды, морские огурцы, хрупкие звезды, и морские лилии. Как и другие иглокожие, они обладают пятиступенчатой симметрией (называемой пентамеризм ) и перемещаться с помощью сотен крошечных прозрачных клейких материалов "трубка ноги ". Симметрия не очевидна у живого животного, но хорошо видна в высушенном тест.[2]
В частности, термин «морской еж» относится к «обычным ехиноидам», которые являются симметричными и шаровидными, и включает несколько различных таксономических групп с двумя подклассами: Euechinoidea («современные» морские ежи, в том числе и нерегулярные) и Cidaroidea или «грифельные ежи» с очень толстыми тупыми шипами, на которых растут водоросли и губки. «Нерегулярные» морские ежи - это инфра-класс внутри Euechinoidea, называемый Irregularia, и включать Ателостомы и Неогнатостомы. Неправильные ехиноиды включают: уплощенные песочные доллары, морское печенье, и сердечные ежи.[3]
Вместе с морскими огурцами (Holothuroidea ), они составляют подтип Echinozoa, который характеризуется шаровидной формой без рукавов и выступающих лучей. Морские огурцы и неправильные ехиноиды вторично развили разнообразные формы. Хотя многие морские огурцы разветвились щупальца окружающие их ротовые отверстия, они произошли от модифицированных трубчатых ножек и не гомологичны рукавам лилий, морских звезд и хрупких звезд.[2]

Paracentrotus lividus, обычный морской еж (Euechinoidea, инфракласс Каринацея )

А песчаный доллар, нерегулярный морской еж (Irregularia )

Phyllacanthus imperialis, цидароидный морской еж (Cidaroidea )

Черный морской еж взят с острова Агиранган, Маангас, Презенсион, Камаринес-Сур, Филиппины.



Описание

Ежи обычно имеют размер от 3 до 10 см (от 1 до 4 дюймов), хотя самые крупные виды могут достигать 36 см (14 дюймов).[4] У них жесткое, обычно сферическое тело, несущее подвижные шипы, что дает учебный класс название Echinoidea (от греч. эхинос, позвоночник).[5] Название «еж» - старое слово для обозначения Ежик, на которые похожи морские ежи; их архаично называли морские ежики.[6][7] Название происходит от старофранцузского Herichun, от латинского Эриций, Ежик.[8]
Как и у других иглокожих, ранние личинки морского ежа обладают двусторонней симметрией,[9] но по мере взросления они развивают пятикратную симметрию. Это наиболее очевидно у «обычных» морских ежей, которые имеют тела примерно сферической формы с пятью частями одинакового размера, расходящимися от их центральных осей. Рот находится у основания животного, а задний проход - вверху; нижняя поверхность описывается как «оральная», а верхняя - как «аборальная».[а][2]
Однако некоторые морские ежи, в том числе ежи, имеют овальную форму с четкими передними и задними концами, что придает им степень двусторонней симметрии. У этих ежей верхняя поверхность тела слегка выпуклая, но нижняя сторона плоская, а по бокам отсутствуют трубчатые ножки. Эта «неправильная» форма тела эволюционировала, чтобы позволить животным рыться в песке или других мягких материалах.[4]
Системы
Скелетно-мышечный

Морские ежи могут показаться неспособен двигаться но это ложное впечатление. Иногда наиболее заметным признаком жизни являются шипы, которые прикреплены к шарнирным соединениям и могут указывать в любом направлении; у большинства ежей прикосновение вызывает быструю реакцию шипов, которые сходятся к точке прикосновения. У морских ежей нет видимых глаз, ног или средств передвижения, но они могут свободно, но медленно перемещаться по твердым поверхностям, используя липкие трубчатые ножки, работая вместе с шипами.[2]
Внутренние органы заключены в твердую оболочку или тест, состоящий из сросшихся пластин карбонат кальция покрытый тонким дерма и эпидермис. Тест жесткий, делится на пять амбулакральных бороздок, разделенных пятью межмбулакральными областями. Каждая из этих областей состоит из двух рядов пластин, поэтому в тесте на морских ежах всего 20 рядов пластин. Пластинки покрыты округлыми бугорками, в которых расположены гнезда, к которым прикреплены иглы. шаровые и шарнирные соединения. Внутренняя поверхность теста облицована брюшина.[4] Морские ежи превращают водянистую углекислый газ используя каталитический процесс с участием никель в часть теста с карбонатом кальция.[10]
У большинства видов есть две серии шипов: первичная (длинная) и вторичная (короткая), распределенная по поверхности тела, с самой короткой на полюсах и самой длинной на экваторе. Шипы обычно полые и цилиндрические. Сокращение мышечной оболочки, покрывающей тест, заставляет позвоночник наклоняться в том или ином направлении, в то время как внутренняя оболочка коллагеновых волокон может обратимо изменяться с мягкой на жесткую, что может зафиксировать позвоночник в одном положении. Между колючками расположено несколько типов педицеллярия, подвижные стебельчатые конструкции с челюстями.[2]
Морские ежи передвигаются, используя свои многочисленные гибкие трубчатые ножки, подобно морским звездам; У обычных морских ежей нет любимого направления ходьбы.[11] Ножки трубки выступают через пары пор в тесте и управляются водная сосудистая система; это работает через гидравлическое давление, позволяя морскому ежу закачивать воду в ножки трубки и из них. Во время передвижения ножкам трубки помогают шипы, которые можно использовать для подталкивания тела или подъема теста с подложки. Движение обычно связано с кормлением, при этом красный морской еж (Mesocentrotus franciscanus) примерно 7,5 см (3 дюйма) в день, когда есть достаточно еды, и до 50 см (20 дюймов) в день, когда ее нет. Перевернутый морской еж может выпрямиться, постепенно прикрепляя и отсоединяя свои трубчатые лапы и манипулируя своими шипами, чтобы повернуть тело вертикально.[2] Некоторые виды зарываются в мягкий осадок, используя свои шипы, и Paracentrotus lividus использует свои челюсти, чтобы зарываться в мягкие камни.[12]

Испытание Echinus esculentus, обычный морской еж

Испытание Echinodiscus tenuissimus, нерегулярный морской еж ("песчаный доллар ")

Испытание Phyllacanthus imperialis, а цидароид морской еж. Для них характерны большие бугорки с крупными радиолами.

Крупный план теста, показывающий амбулакральную бороздку с двумя рядами пар пор между двумя межмбулакральными областями (зеленый). Бугорки неперфорированные.

Крупный план апикального диска цидароидного морского ежа: 5 отверстий - гонопоры, а центральное - анус («перипрокт»). Самая большая генитальная пластинка - это мадрепорит.[13]




Питание и пищеварение

Рот находится в центре поверхности рта у обычных ежей или ближе к одному концу у нерегулярных ежей. Он окружен губами из более мягкой ткани с многочисленными небольшими вкрапленными костями. Эта область, называемая перистомом, также включает пять пар модифицированных трубчатых ножек и, у многих видов, пять пар жабр.[4] Челюстной аппарат состоит из пяти прочных пластин в форме стрелы, известных как пирамиды, на вентральной поверхности каждой из которых имеется зубчатый венец с твердым зубом, направленным к центру рта. Специализированные мышцы контролируют выступание аппарата и действие зубов, и животное может хватать, царапать, тянуть и рвать.[2] Структура рта и зубов оказалась настолько эффективной при захвате и измельчении, что аналогичные структуры были протестированы для использования в реальных условиях.[14]
На верхней поверхности теста у аборального полюса находится перепонка, перипрокт, который окружает анус. Перипрокт содержит различное количество твердых пластин, пять из которых, генитальные пластины, содержат гонопоры, а одна модифицирована, чтобы содержать мадрепорит, который используется для балансировки водной сосудистой системы.[2]

Рот большинства морских ежей состоит из пяти зубов или пластин из карбоната кальция, внутри которых имеется мясистая язычковая структура. Весь жевательный орган известен как фонарь Аристотеля из Аристотель описание в его История животных.
- ... у ежа есть то, что мы в основном называем головой и ртом внизу, и место для выхода остатков вверху. У ежа также есть пять полых зубов внутри, а в середине этих зубов мясистая субстанция, служащая служению язык. Рядом с этим идет пищевод, а затем желудок, разделенный на пять частей и заполненный экскрементами, все пять частей соединяются в анальный отверстие, где раковина перфорирована для выхода ... В действительности ротовой аппарат ежа сплошной от одного конца до другого, но внешне это не так, но выглядит как рог фонарь с опущенными стеклами рога. (Тр. Д'Арси Томпсон )
Однако недавно было доказано, что это неправильный перевод. Фонарь Аристотеля на самом деле относится ко всей форме морских ежей, которые выглядят как древние лампы времен Аристотеля.[15][16]
Сердечные ежи необычны тем, что не имеют фонаря. Вместо этого рот окружен реснички которые тянут полоски слизи, содержащей частицы пищи, к ряду бороздок вокруг рта.[4]
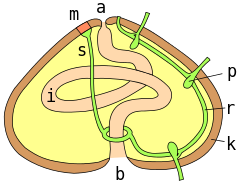
а = анус ; m = мадрепорит ; s = канал водоносного горизонта; r = радиальный канал; p = подиальная ампула; k = тестовая стена; я = кишечник ; b = рот
Фонарь, если он есть, окружает ротовую полость и глотка. В верхней части фонаря глотка открывается в пищевод, который спускается вниз по внешней стороне фонаря, чтобы присоединиться к маленькому кишечник и один слепая кишка. Тонкая кишка проходит полный круг вокруг внутренней части теста, прежде чем присоединиться к толстой кишке, которая завершает еще один цикл в противоположном направлении. Из толстой кишки прямая кишка поднимается к анусу. Несмотря на названия, тонкий и толстый кишечник морских ежей никоим образом не гомологичный к одноименным структурам у позвоночных.[4]
Пищеварение происходит в кишечнике, а слепая кишка производит дальнейшее пищеварение. ферменты. Дополнительная трубка, называемая сифоном, проходит рядом с большей частью кишечника, открываясь в него с обоих концов. Может участвовать в резорбции воды из пищи.[4]
Кровообращение и дыхание
Водяная сосудистая система ведет вниз от мадрепорита через тонкий каменный канал к кольцевому каналу, окружающему пищевод. Отсюда через каждую амбулакральную область проходят радиальные каналы, которые заканчиваются маленьким щупальцем, которое проходит через амбулакральную пластинку около аборального полюса. От этих радиальных каналов отходят боковые каналы, заканчиваясь ампулами. Отсюда две трубки проходят через пару пор на пластине и заканчиваются ножками трубки.[2]
Морские ежи обладают гемальной системой со сложной сетью сосудов в брыжейках вокруг кишечника, но о функционировании этой системы известно немного.[2] Однако основная циркулирующая жидкость заполняет общую полость тела или целомудрие. Эта целомическая жидкость содержит фагоцитарный целомоциты, которые перемещаются по сосудистой и гемальной системам и участвуют во внутреннем транспорте и газообмене. Целомоциты являются важной частью свертывание крови, но также собирают продукты жизнедеятельности и активно выводят их из организма через жабры и трубочные ножки.[4]
Большинство морских ежей имеют пять пар наружных жабр, прикрепленных к перистомиальной мембране вокруг рта. Эти тонкостенные выступы полости тела являются основными органами дыхания тех ежей, которые ими обладают. Жидкость может перекачиваться через внутреннюю часть жабр с помощью мышц, связанных с фонарем, но это не обеспечивает непрерывного потока, а происходит только тогда, когда у животного мало кислорода. Трубчатые ножки также могут действовать как органы дыхания и являются основными местами газообмена у сердечных ежей и морских ежей, у которых отсутствуют жабры. Внутренняя часть каждой ножки трубки разделена перегородкой, которая уменьшает диффузию между входящим и выходящим потоками жидкости.[2]

Нервная система и чувства
Нервная система морских ежей имеет относительно простое устройство. Без настоящего мозга нервный центр представляет собой большое нервное кольцо, окружающее рот внутри фонаря. От нервного кольца пять нервов исходят под лучевыми каналами водной сосудистой системы и разветвляются на многочисленные более тонкие нервы, иннервирующие ножки трубки, позвоночник и педицеллярии.[4]
Морские ежи чувствительны к прикосновениям, свету и химическим веществам. В эпителии имеется множество чувствительных клеток, особенно в шипах, педицеллярии и трубчатых ножках, а также вокруг рта.[2] Хотя у них нет глаз или глазных пятен (кроме диадемы, которые могут следовать за угрозой своими шипами), все тело большинства обычных морских ежей может функционировать как сложный глаз.[17] Обычно морских ежей негативно привлекает свет, и они стремятся спрятаться в расщелинах или под предметами. Большинство видов, кроме карандаш ежи, имеют статоцисты в шаровых органах, называемых сферидиями. Это стеблевые структуры, расположенные в амбулакральных областях; их функция - помогать в гравитационной ориентации.[4]
История жизни
Размножение

Морские ежи раздельнополый, имея отдельные мужской и женский пол, хотя внешне не видно никаких отличительных черт. Помимо их роли в воспроизводстве, гонады также являются органами хранения питательных веществ и состоят из двух основных типов клеток: стволовые клетки, и соматические клетки называемые питательными фагоцитами.[18] Обычные морские ежи имеют пять гонад, лежащих под межмбулакральными областями раковины, в то время как неправильные формы в основном имеют четыре гонады, причем задние гонады отсутствуют; Сердечных ежей их три-два. Каждая гонада имеет единственный проток, поднимающийся от верхнего полюса и открывающийся в гонопора лежит в одной из половых пластин, окружающих задний проход. Некоторые роющие ежи имеют удлиненный сосочек, который позволяет высвобождать гаметы над поверхностью осадка.[2] Гонады выстланы мускулами под брюшиной, что позволяет животному сжимать гаметы через канал в окружающую морскую воду, где оплодотворение происходит.[4]
Разработка

В период раннего развития морской еж эмбрион проходит 10 циклов деление клеток,[19] в результате чего один эпителиальный слой, охватывающий бластоцель. Затем зародыш начинает гаструляция, многокомпонентный процесс, который резко меняет свою структуру за счет инвагинация произвести три ростковые отростки с участием эпителиально-мезенхимальный переход; начальный мезенхима клетки переходят в бластоцель[20] и стать мезодерма.[21] Было высказано предположение, что эпителиальная полярность вместе с плоской клеточной полярностью может быть достаточно, чтобы управлять гаструляцией у морского ежа.[22]

Необычной особенностью развития морского ежа является замена личинки двусторонняя симметрия пятеричной симметрией взрослого. Во время дробления уточняются мезодерма и мелкие микромеры. В конце гаструляции клетки этих двух типов образуют целомический мешочки. На личиночных стадиях имаго зачаток вырастает из левого целомического мешка; после метаморфозы этот зачаток вырастает и становится взрослым. В животно-растительная ось устанавливается до оплодотворения яйца. Ось орально-аборальная определяется на ранней стадии дробления, а ось влево-вправо появляется на стадии поздней гаструлы.[23]
Жизненный цикл и развитие

В большинстве случаев яйца самок свободно плавают в море, но некоторые виды держатся за них шипами, обеспечивая большую степень защиты. Неоплодотворенная яйцеклетка встречается со свободно плавающей спермой, выпущенной самцами, и превращается в свободно плавающую бластула эмбрион всего за 12 часов. Первоначально простой клубок клеток, вскоре бластула трансформирует в конус эхиноплютеус личинка. У большинства видов эта личинка имеет 12 удлиненных плеч, выстланных полосами ресничек, которые захватывают частицы пищи и переносят их ко рту. У некоторых видов бластула содержит запасы питательных веществ. желток у него нет оружия, потому что его не нужно кормить.[4]
Для завершения развития личинки требуется несколько месяцев, переход во взрослую форму начинается с образования тестовых пластинок в ювенильном зачатке, который развивается с левой стороны личинки, причем его ось перпендикулярна оси личинки. Вскоре личинка опускается на дно и метаморфозы в молодого ежа всего за час.[2] У некоторых видов взрослые особи достигают максимального размера примерно за пять лет.[4] В фиолетовый еж становится половозрелым через два года и может прожить двадцать.[24]
Экология
Трофический уровень

Питаются морские ежи в основном водоросли, поэтому они в первую очередь травоядные животные, но может питаться морскими огурцами и различными беспозвоночными, такими как моллюски, полихеты, губки, хрупкие звезды и морские лилии, что делает их всеядными, потребителями в различных трофические уровни.[25]
Хищники, паразиты и болезни
О массовой смертности морских ежей впервые сообщили в 1970-х годах, но до появления аквакультуры болезни морских ежей были мало изучены. В 1981 году бактериальная «пятнистость» вызвала почти полную смертность молодых особей. Псевдоцентротус депрессивный и Hemicentrotus pulcherrimus оба культивируются в Японии; заболевание рецидивировало в последующие годы. Она подразделялась на холодноводную «родниковую» болезнь и горячую «летнюю» форму.[26] Другое условие, болезнь лысого морского ежа, вызывает потерю шипов и поражения кожи и, как полагают, имеет бактериальное происхождение.[27]
Взрослые морские ежи обычно хорошо защищены от большинства хищников своими сильными и острыми шипами, которые у некоторых видов могут быть ядовитыми.[28] Маленький морской еж живет среди колючек ежей, таких как Диадема; молодые особи питаются педицелляриями и сферидиями, взрослые самцы выбирают трубчатые лапы, а взрослые самки уходят, чтобы питаться яйцами креветок и моллюсками.[29]
Морские ежи - одно из любимых блюд многих лобстеры, крабы, спинорог, Калифорнийская овчарка, морская выдра и волк угорь (которые специализируются на морских ежах). Все эти животные обладают особыми приспособлениями (зубы, клешни, когти) и силой, которые позволяют им преодолевать превосходные защитные свойства морских ежей. Не обращая внимания на хищников, ежи разрушают свою среду обитания, создавая то, что биологи называют бесплодный еж, лишенный макроводорослей и связанных фауна.[30] Морские выдры снова вошли британская Колумбия, резко улучшая здоровье прибрежных экосистем.[31]

Волчий угорь, узкоспециализированный хищник морских ежей

А морская выдра питаясь фиолетовый морской еж.

Краб (Carpilius convxus ) нападение грифельного карандаша морского ежа (Heterocentrotus mamillatus )

А губан отделка останков поврежденного Tripneustes gratilla




Защита от хищников

В шипы длинные и острые у некоторых видов, защищают ежа от хищники. Некоторые тропические морские ежи любят Diadematidae, Echinothuriidae и Toxopneustidae имеют ядовитые шипы. Другие существа также используют эту защиту; крабы, креветки и другие организмы укрываются среди колючек и часто принимают окраску своего хозяина. Некоторые крабы в Dorippidae семья носит в когтях морских ежей, морских звезд, острые панцири или другие защитные предметы.[32]
Педицелларии[33] являются хорошим средством защиты от эктопаразитов, но не панацеей, поскольку некоторые из них фактически питаются им.[34] Гемальная система защищает от эндопаразитов.[35]
Диапазон и среда обитания
Морские ежи обитают в большинстве мест обитания на морском дне, начиная с приливной зоны и ниже, на чрезвычайно широком диапазоне глубин.[36] Некоторые виды, такие как Cidaris Abyssicola, могут жить на глубине до нескольких тысяч метров. Многие роды встречаются только в абиссальная зона, в том числе многие цидароиды, большинство родов в Echinothuriidae семья, и «кактусовые ежи» Дермехинус. Одна из самых обеспеченных семей - Pourtalesiidae,[37] странные морские ежи неправильной формы в форме бутылки, которые живут только в хадальная зона и были собраны на глубине 6850 метров под поверхностью в Зондский желоб.[38] Тем не менее, это делает морских ежей классом иглокожих, живущих наименее глубоко по сравнению с хрупкие звезды, морская звезда и морские лилии которые остаются обильными ниже 8000 м (26 250 футов) и морские огурцы которые были зарегистрированы с высоты 10 687 м (35 100 футов).[38]
Плотность населения варьируется в зависимости от среды обитания, с более плотным населением в бесплодных районах по сравнению с ламинария стоит.[39][40] Даже в этих бесплодных районах наибольшая плотность наблюдается на мелководье. Популяции обычно обитают в более глубоких водах при наличии волн.[40] Плотность уменьшается зимой, когда штормы заставляют их искать защиту в трещинах и вокруг более крупных подводных сооружений.[40]В галечный еж (Colobocentrotus atratus), обитающий на открытых берегах, особенно устойчив к воздействию волн. Это один из немногих морских ежей, которые могут выдерживать много часов без воды.[41]
Морских ежей можно встретить в любом климате, от теплых морей до полярных океанов.[36] Личинки полярного морского ежа Sterechinus neumayeri было обнаружено, что энергия используется в обменных процессах в двадцать пять раз эффективнее, чем у большинства других организмов.[42] Несмотря на то, что они присутствуют почти во всех морских экосистемах, большинство видов обитает на побережьях умеренного и тропического климата, между поверхностью и глубиной нескольких десятков метров, недалеко от моря. фотосинтетический Источники питания.[36]

Фиолетовые морские ежи во время отлива Калифорния. Они роют в скале углубление, чтобы днем укрыться от хищников.

Dermechinus horridus, абиссальный вид, на глубине тысяч метров

Антарктический морской еж (Sterechinus neumayeri ) обитает в замерзших морях.

Форма галечный еж позволяет ему оставаться на забитых волнами скалах.




Эволюция
История окаменелостей

Самые ранние окаменелости ехиноидов относятся к верхней части Ордовик период (около 450 Mya ).[43] Существует богатая летопись окаменелостей, их тяжелые испытания сделаны из кальцит плиты, сохранившиеся в скалах с тех пор.[44]Колючки присутствуют у некоторых хорошо сохранившихся экземпляров, но обычно остается только раковина. Изолированные шипы встречаются в окаменелостях. Немного Юрский и Меловой Cidaroida были очень тяжелые, булавовидные шипы.[45]
Наиболее ископаемое ехиноиды из Палеозой эры неполные, состоящие из отдельных шипов и небольших скоплений разрозненных пластин раздавленных особей, в основном в Девонский и Каменноугольный горные породы. Мелководье известняки от ордовика и Силурийский периоды Эстония славятся ехиноидами.[46] Палеозойские ежи, вероятно, населяли относительно спокойные воды. Из-за их тонких панцирей они, конечно, не выжили бы в заброшенных волнами прибрежных водах, населенных многими современными ехиноидами.[46] Ехиноиды почти исчезли в конце палеозойской эры. Пермский период период. Только две линии пережили массовое исчезновение этого периода и попали в Триасовый: род Миоцидарис, которая дала начало современным цидароида (ежи карандаш), и предок, который дал начало эуэхиноиды. К верхнему триасу их количество снова увеличилось. Цидароиды очень мало изменились с позднего Триасовый, и являются единственной сохранившейся палеозойской группой ехиноидов.[46]
Евехиноиды разошлись в новые линии в Юрский и Меловой периоды, и из них появились первые неправильные ехиноиды ( Ателостомы ) в ранней юре.[47]
Некоторые ехиноиды, например Микрастер в мелу мелового периода служат зоной или индекс окаменелости. Поскольку их много и они быстро эволюционируют, они позволяют геологам датировать окружающие породы.[48]
в Палеоген и Неоген периоды (около От 66 до 1,8 млн лет назад), песочные доллары (Clypeasteroida). Их характерные плоские раковины и крошечные иглы были приспособлены к жизни на рыхлом песке или под ним на мелководье, и они широко представлены в виде окаменелостей в южноевропейских известняках и песчаниках.[46]

Археоцидарис Brownwoodensis, Cidaroida, Каменноугольный, c. 300 млн лет назад

Миоцидарис Коаева, Cidaroida, Середина Триасовый, c. 240 млн лет назад

Наличник плотти, Irregularia, Середина Юрский, c. 162 млн лет назад

Эхинокорис, Holasteroida, Верхний Меловой, c. 80 млн лет назад

Эхинолампы овальный, Кассидулоида, Середина эоцен, c. 40 млн лет назад

Clypeaster предвестник, Клипеастероиды, Миоцен, c. 10 млн лет






Филогения
Внешний
Ехиноиды дейтеростом животные, как хордовые. Анализ 219 генов всех классов иглокожих в 2014 г. дает следующее филогенетическое дерево.[49] Приблизительные даты ветвления основных клад указаны в миллионах лет назад (млн лет назад).
| Bilateria |
| ||||||||||||||||||||||||||||||||||||||||||||||||





Внутренний
Филогения морских ежей следующая:[50][51]
| Echinoidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 450 млн лет назад |

Отношение к людям
Травмы
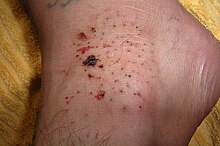
Травмы морского ежа - это колотые раны, нанесенные хрупкими хрупкими шипами животного.[52]Они являются частым источником травм океанских пловцов, особенно на прибрежных поверхностях, где обитают кораллы с неподвижными морскими ежами. Их укусы различаются по силе в зависимости от вида. Их шипы могут быть ядовитыми или вызывать инфекцию. Гранулема также может произойти окрашивание кожи естественным красителем внутри морского ежа. Проблемы с дыханием могут указывать на серьезную реакцию морского ежа на токсины.[53] Они наносят болезненную рану, когда проникают через кожу человека, но сами по себе не опасны, если полностью удалить их сразу; если его оставить на коже, могут возникнуть дальнейшие проблемы.[54]
Наука
Морские ежи традиционные модельные организмы в биология развития. Это использование возникло в 1800-х годах, когда их эмбриональное развитие стало легко просматриваться под микроскопом. Прозрачность яиц ежа позволила использовать их для наблюдения сперма клетки действительно оплодотворяют яйцеклетка.[55] Их продолжают использовать для эмбриональных исследований, так как пренатальное развитие продолжает искать тесты на смертельные заболевания. Морские ежи используются в исследованиях долголетия для сравнения молодых и старых представителей этого вида, особенно из-за их способности при необходимости регенерировать ткани.[56] Ученые из Сент-Эндрюсский университет обнаружили генетическую последовательность, область 2А, у морских ежей, которые ранее считались принадлежащими только вирусам, поражающим людей, таким как вирус ящура.[57]В последнее время, Эрик Х. Дэвидсон и Рой Джон Бриттен выступали за использование ежей в качестве модельных организмов из-за их легкой доступности, высокой плодовитости и долгой продолжительности жизни. Вне эмбриология, ежи дают возможность исследовать цис-регуляторные элементы.[58]Океанография проявляла интерес к мониторингу здоровья ежей и их популяций как способу общей оценки закисление океана,[59] температуры и экологические воздействия.
Эволюционное расположение организма и уникальная эмбриология с пятикратной симметрией были главными аргументами в предложении попытаться установить последовательность его развития. геном. Важно отметить, что ежи действуют как ближайшие живые существа по отношению к хордовым, и поэтому представляют интерес тем, что они могут пролить свет на эволюцию позвоночные.[60] Геном Стронгилоцентротус пурпуратус, был завершен в 2006 году и установил гомологию между морскими ежами и позвоночными. иммунная система -связанные гены. Код морских ежей не менее 222 Толл-подобный рецептор гены и более 200 генов, связанных с Nod-подобный рецептор семейство позвоночных.[61] Это увеличивает его полезность как ценный модельный организм для изучения эволюция из врожденный иммунитет. Секвенирование также показало, что, хотя считалось, что некоторые гены относятся только к позвоночным, были также инновации, которые ранее никогда не встречались за пределами классификации хордовых, такие как факторы иммунной транскрипции. PU.1 и СПИБ.[60]
Как еда
Гонады как самцов, так и самок морских ежей, обычно называемых икрой морского ежа или кораллами,[62] являются кулинарными деликатесами во многих частях мира.[63][64][65]В Средиземноморская кухня, Paracentrotus lividus часто едят сырым или с лимоном,[66] и известный как Риччи на Итальянский меню, где он иногда используется в соусах для пасты. Он также может ароматизировать омлеты, яичница-болтунья, рыбный суп,[67] майонез, соус бешамель для тарталеток,[68] то були для суфле,[69] или же Голландский соус приготовить рыбный соус.[70] В Чилийская кухня подается сырым с лимоном, луком и оливковым маслом. Хотя съедобный Strongylocentrotus droebachiensis Водится в Северной Атлантике, в пищу его мало. Однако морские ежи (называемые Уутук в Алутик ) обычно едят коренное население Аляски около Остров Кадьяк. Обычно он экспортируется, в основном в Япония.[71]В Вест-Индии грифельный карандаш ежи съедены.[63]На тихоокеанском побережье Северной Америки, Strongylocentrotus franciscanus получил похвалу от Оуэль Гиббонс; Стронгилоцентротус пурпуратус тоже едят.[63]В Новой Зеландии Evechinus chloroticus, известный как кина в маори, это деликатес, который традиционно едят в сыром виде. Хотя новозеландские рыбаки хотели бы экспортировать их в Японию, их качество слишком непостоянно.[72]В Японии морского ежа называют уни (う に)и его икра продается по цене 40 000 иен (360 долларов США) за кг;[73] он подается в сыром виде сашими или в суши, с соевый соус и васаби. Япония в больших количествах импортирует из США, Южная Корея, и другие производители. Япония потребляет 50 000 тонн ежегодно, что составляет более 80% мирового производства.[74] Спрос на морских ежей в Японии вызывает опасения по поводу чрезмерного вылова рыбы.[75]Коренные американцы в Калифорнии также известны морские ежи.[76] Побережье Южной Калифорнии известно как источник высококачественной рыбы, и дайверы собирают морских ежей из зарослей водорослей на глубинах до 24 м / 80 футов.[77] По состоянию на 2013 год государство ограничивало практику 300 лицензиями дайвера на морских ежей.[77]

Японский uni-ikura don, яйцо морского ежа и яйцо лосося домбури

Открытые морские ежи на Сицилии



Аквариум

Некоторые виды морских ежей, например, грифельный еж (Eucidaris tribuloides ), обычно продаются в аквариумных магазинах. Некоторые виды эффективно контролируют нитчатые водоросли, и они хорошо дополняют беспозвоночный бак.[79]
Фольклор
Народная традиция в Дании и южной Англии считала окаменелости морских ежей молниями, способными отражать вред от молнии или колдовства, как апотропный символ.[80] Другая версия предполагала, что это были окаменевшие яйца змей, способные защитить от болезней сердца и печени, ядов и травм в бою, и, соответственно, их несли как амулеты. Согласно легенде, они были созданы магией из пены, сделанной змеями в разгар лета.[81]
Смотрите также
Примечания
Рекомендации
- ^ "Сеть разнообразия животных - Echinoidea". Музей зоологии Мичиганского университета. Получено 26 августа 2012.
- ^ а б c d е ж грамм час я j k л м п о Рупперт, Эдвард Э .; Фокс, Ричард, С .; Барнс, Роберт Д. (2004). Зоология беспозвоночных, 7-е издание. Cengage Learning. С. 896–906. ISBN 978-81-315-0104-7.
- ^ Kroh, A .; Ханссон, Х. (2013). "Echinoidea (Леске, 1778 г.) ". Черви. Всемирный регистр морских видов. Получено 2014-01-04.CS1 maint: несколько имен: список авторов (связь)
- ^ а б c d е ж грамм час я j k л м Барнс, Роберт Д. (1982). Зоология беспозвоночных. Филадельфия, Пенсильвания: Holt-Saunders International. С. 961–981. ISBN 0-03-056747-5.
- ^ Гилл, Майкл. "Таксономические этимологии EEOB 111". Получено 13 марта 2018.
- ^ Райт, Энн. 1851 г. Наблюдающий глаз или письма детям о трех низших сферах жизни животных. Лондон: Джарролд и сыновья, стр. 107.
- ^ Сойер, Алексис. 1853. Пантрофеон или История еды и ее приготовления: с самых ранних веков мира. Бостон: Тикнор, Рид и Филдс, стр. 245.
- ^ "еж (сущ.)". Интернет-словарь этимологии. Получено 13 марта 2018.
- ^ Стахан и Рид, Молекулярная генетика человека, п. 381: "Что делает нас людьми"
- ^ «Морские ежи открывают многообещающую альтернативу улавливанию углерода». Гизмаг. 4 февраля 2013 г.. Получено 2013-02-05.
- ^ Казуя Ёсимура, Томоаки Икетани и Тацуо Мотокава, «Отдают ли обычные морские ежи предпочтение, в какой части тела они ориентируются вперед при ходьбе?», Морская биология, т. 159, № 5, 2012 г., с. 959–965.
- ^ Boudouresque, Charles F .; Верлак, Марк (2006). "13: Экология Paracentrotus lividus". В Лоуренс, Джон, М. (ред.). Съедобные морские ежи: биология и экология. Эльзевир. п. 243. ISBN 978-0-08-046558-6.
- ^ «Апикальный диск и перипрокт». Музей естественной истории, Лондон. Получено 2 ноября 2019.
- ^ Коготь, вдохновленный ртом морских ежей, может выкапывать марсианский грунт
- ^ Вултсиаду, Элени; Чинтироглу, Харитон (2008). «Фонарь Аристотеля в иглокожих: древняя загадка». Cahiers de Biologie Marine. Биологическая станция Роскофф. 49 (3): 299–302.[постоянная мертвая ссылка ]

- ^ Чой, Чарльз К. (29 декабря 2010 г.). «У морских ежей-камнеедов зубы самозатачивающиеся». Новости National Geographic. Получено 2017-11-12.
- ^ Найт, К. (2009). «Морские ежи используют все тело как глаз». Журнал экспериментальной биологии. 213 (2): i – ii. Дои:10.1242 / jeb.041715. Сложить резюме – LiveScience (28 декабря 2009 г.).
- ^ Gaitán-Espitia, J.D .; Sánchez, R .; Bruning, P .; Карденас, Л. (2016). «Функциональное понимание транскриптома семенников съедобного морского ежа Loxechinus albus». Научные отчеты. 6: 36516. Bibcode:2016НатСР ... 636516Г. Дои:10.1038 / srep36516. ЧВК 5090362. PMID 27805042.
- ^ А. Гайон, А. Скудери; Д. Пеллегрини; Д. Сартори (2013). «Воздействие мышьяка влияет на развитие зародыша морского ежа Paracentrotus lividus (Lamarck, 1816)». Бюллетень загрязнения окружающей среды и токсикологии. 39 (2): 124–8. Дои:10.3109/01480545.2015.1041602. PMID 25945412. S2CID 207437380.
- ^ Коминами, Тэцуя; Таката, Хироми (2004). «Гаструляция у эмбриона морского ежа: модельная система для анализа морфогенеза однослойного эпителия». Развитие, рост и дифференциация. 46 (4): 309–26. Дои:10.1111 / j.1440-169x.2004.00755.x. PMID 15367199.
- ^ Shook, D; Келлер, Р. (2003). «Механизмы, механика и функция эпителиально-мезенхимальных переходов в раннем развитии». Механизмы развития. 120 (11): 1351–83. Дои:10.1016 / j.mod.2003.06.005. PMID 14623443. S2CID 15509972.; Катов, Хидеки; Солурш, Майкл (1980). «Ультраструктура проникновения клеток первичной мезенхимы у морского ежа Lytechinus pictus». Журнал экспериментальной зоологии. 213 (2): 231–246. Дои:10.1002 / jez.1402130211.; Балинский, Б.И. (1959). «Электромикроскопическое исследование механизмов адгезии клеток в бластуле и гаструле морского ежа». Экспериментальные исследования клеток. 16 (2): 429–33. Дои:10.1016/0014-4827(59)90275-7. PMID 13653007.; Hertzler, PL; Макклей Д.Р. (1999). «alphaSU2, эпителиальный интегрин, который связывает ламинин в эмбрионе морского ежа». Биология развития. 207 (1): 1–13. Дои:10.1006 / dbio.1998.9165. PMID 10049560.; Финк, РД; Макклей Д.Р. (1985). "Three cell recognition changes accompany the ingression of sea urchin primary mesenchyme cells". Биология развития. 107 (1): 66–74. Дои:10.1016/0012-1606(85)90376-8. PMID 2578117.; Burdsal, CA; Alliegro, MC; McClay, DR (1991). "Tissue-specific, temporal changes in cell adhesion to echinonectin in the sea urchin embryo". Биология развития. 144 (2): 327–34. Дои:10.1016/0012-1606(91)90425-3. PMID 1707016.; Miller, JR; McClay, DR (1997). "Characterization of the Role of Cadherin in Regulating Cell Adhesion during Sea Urchin Development". Биология развития. 192 (2): 323–39. Дои:10.1006/dbio.1997.8740. PMID 9441671.; Miller, JR; McClay, DR (1997). "Changes in the pattern of adherens junction-associated beta-catenin accompany morphogenesis in the sea urchin embryo". Биология развития. 192 (2): 310–22. Дои:10.1006/dbio.1997.8739. PMID 9441670.; Anstrom, JA (1989). "Sea urchin primary mesenchyme cells: ingression occurs independent of microtubules". Биология развития. 131 (1): 269–75. Дои:10.1016/S0012-1606(89)80058-2. PMID 2562830.; Anstrom, JA (1992). "Microfilaments, cell shape changes, and the formation of primary mesenchyme in sea urchin embryos". Журнал экспериментальной зоологии. 264 (3): 312–22. Дои:10.1002/jez.1402640310. PMID 1358997.
- ^ Nissen, Silas Boye; Rønhild, Steven; Trusina, Ala; Sneppen, Kim (November 27, 2018). "Theoretical tool bridging cell polarities with development of robust morphologies". eLife. 7: e38407. Дои:10.7554/eLife.38407. ЧВК 6286147. PMID 30477635.
- ^ Warner, Jacob F.; Lyons, Deirdre C.; McClay, David R. (2012). "Left-Right Asymmetry in the Sea Urchin Embryo: BMP and the Asymmetrical Origins of the Adult". PLOS Биология. 10 (10): e1001404. Дои:10.1371/journal.pbio.1001404. ЧВК 3467244. PMID 23055829.
- ^ Worley, Alisa (2001). "Стронгилоцентротус пурпуратус". Сеть разнообразия животных. Получено 2016-12-05.
- ^ Baumiller, Tomasz K. (2008). "Crinoid Ecological Morphology". Ежегодный обзор наук о Земле и планетах. 36: 221–49. Bibcode:2008AREPS..36..221B. Дои:10.1146/annurev.earth.36.031207.124116.
- ^ Lawrence, John M. (2006). Edible Sea Urchins: Biology and Ecology. Эльзевир. С. 167–168. ISBN 978-0-08-046558-6.
- ^ Jangoux, Michel (1987). "Diseases of Echinodermata. I. Agents microorganisms and protistans". Болезни водных организмов. 2: 147–162. Дои:10.3354/dao002147.
- ^ "Defence – spines". Echinoid Directory. Музей естественной истории.
- ^ Sakashita, Hiroko (1992). "Sexual dimorphism and food habits of the clingfish, Diademichthys lineatus, and its dependence on host sea urchin". Экологическая биология рыб. 34 (1): 95–101. Дои:10.1007/BF00004787. S2CID 32656986.
- ^ Terborgh, John; Estes, James A (2013). Trophic Cascades: Predators, Prey, and the Changing Dynamics of Nature. Island Press. п. 38. ISBN 978-1-59726-819-6.
- ^ "Aquatic Species at Risk – Species Profile – Sea Otter". Рыболовство и океаны Канады. Архивировано из оригинал на 2008-01-23. Получено 2007-11-29.
- ^ Тиль, Мартин; Watling, Les (2015). Lifestyles and Feeding Biology. Издательство Оксфордского университета. С. 200–202. ISBN 978-0-19-979702-8.
- ^ "Defence – pedicellariae". Echinoid Directory. Музей естественной истории.
- ^ Hiroko Sakashita, " Sexual dimorphism and food habits of the clingfish, Diademichthys lineatus, and its dependence on host sea urchin ", Environmental Biology of Fishes, vol. 34, no 1, 1994, p. 95–101
- ^ Jangoux, M. (1984). "Diseases of echinoderms" (PDF). Helgoländer Meeresuntersuchungen. 37 (1–4): 207–216. Bibcode:1984HM.....37..207J. Дои:10.1007/BF01989305. S2CID 21863649. Получено 23 марта 2018.
- ^ а б c Kroh, Andreas (2010). «Филогения и классификация постпалеозойских ежей». Журнал систематической палеонтологии. 8 (2): 147–212. Дои:10.1080/14772011003603556..
- ^ Mah, Christopher (April 12, 2011). "Sizes and Species in the Strangest of the Strange : Deep-Sea Pourtalesiid Urchins". The Echinoblog..
- ^ а б Mah, Christopher (8 April 2014). "What are the Deepest known echinoderms?". The Echinoblog. Получено 22 марта 2018..
- ^ Mattison, J.E.; Trent, J.D.; Shanks, AL; Akin, T.B.; Pearse, J.S. (1977). "Movement and feeding activity of red sea urchins (Strongylocentrotus franciscanus) adjacent to a kelp forest". Морская биология. 39 (1): 25–30. Дои:10.1007/BF00395589. S2CID 84338735.
- ^ а б c Konar, Brenda (2000). "Habitat influences on sea urchin populations". В: Hallock and French (Eds). Дайвинг ради науки ... 2000. Материалы 20-го ежегодного научного симпозиума по дайвингу. Американская академия подводных наук. Получено 7 января 2011.
- ^ ChrisM (2008-04-21). "The Echinoblog". echinoblog.blogspot.com.
- ^ Antarctic Sea Urchin Shows Amazing Energy-Efficiency in Nature's Deep Freeze 15 March 2001 Университет Делавэра. Проверено 22 марта 2018 г.
- ^ "Echinoids". Британская геологическая служба. 2017 г.. Получено 14 марта 2018.
- ^ "The Echinoid Directory | Introduction". Музей естественной истории. Получено 16 марта 2018.
- ^ "The Echinoid Directory | Spines". Музей естественной истории. Получено 16 марта 2018.
- ^ а б c d Kirkaldy, J. F. (1967). Fossils in Colour. Лондон: Blandford Press. С. 161–163.
- ^ Schultz, Heinke A.G. (2015). Echinoidea: with pentameral symmetry. Вальтер де Грюйтер. pp. 36 ff, section 2.4. ISBN 978-3-11-038601-1.
- ^ Уэллс, Х.; Хаксли, Джулиан; Wells, G. P. (1931). The Science of life. С. 346–348.
- ^ Telford, M. J.; Lowe, C. J.; Cameron, C.B .; Ortega-Martinez, O.; Aronowicz, J.; Oliveri, P.; Copley, R. R. (2014). "Phylogenomic analysis of echinoderm class relationships supports Asterozoa". Труды Королевского общества B: биологические науки. 281 (1786): 20140479. Дои:10.1098/rspb.2014.0479. ЧВК 4046411. PMID 24850925.
- ^ Planet, Paul J.; Ziegler, Alexander; Schröder, Leif; Ogurreck, Malte; Faber, Cornelius; Stach, Thomas (2012). "Evolution of a Novel Muscle Design in Sea Urchins (Echinodermata: Echinoidea)". PLOS ONE. 7 (5): e37520. Bibcode:2012PLoSO...737520Z. Дои:10.1371/journal.pone.0037520. ЧВК 3356314. PMID 22624043.
- ^ Kroh, Andreas; Smith, Andrew B. (2010). «Филогения и классификация постпалеозойских ежей». Журнал систематической палеонтологии. 8 (2): 147–212. Дои:10.1080/14772011003603556.
- ^ Джеймс, Уильям Д .; Бергер, Тимоти Дж .; и другие. (2006). Кожные болезни Эндрюса: клиническая дерматология. Saunders Elsevier. п. 431. ISBN 0-7216-2921-0.
- ^ Gallagher, Scott A. "Echinoderm Envenomation". eMedicine. Получено 12 октября 2010.
- ^ Matthew D. Gargus; David K. Morohashi (2012). "A sea-urchin spine chilling remedy". Медицинский журнал Новой Англии. 30 (19): 1867–1868. Дои:10.1056/NEJMc1209382. PMID 23134402.
- ^ "Insight from the Sea Urchin". Microscope Imaging Station. Эксплораториум. Архивировано из оригинал на 2017-03-12. Получено 2018-03-07.
- ^ Bodnar, Andrea G.; Coffman, James A. (2016-08-01). "Maintenance of somatic tissue regeneration with age in short- and long-lived species of sea urchins". Aging Cell. 15 (4): 778–787. Дои:10.1111/acel.12487. ISSN 1474-9726. ЧВК 4933669. PMID 27095483.
- ^ Roulston, C.; Luke, G.A.; de Felipe, P.; Ruan, L.; Cope, J.; Nicholson, J.; Sukhodub, A.; Tilsner, J.; Ryan, M.D. (2016). "'2A‐Like' Signal Sequences Mediating Translational Recoding: A Novel Form of Dual Protein Targeting" (PDF). Трафик. 17 (8): 923–39. Дои:10.1111/tra.12411. ЧВК 4981915. PMID 27161495.
- ^ "Sea Urchin Genome Project". sugp.caltech.edu. Архивировано из оригинал на 2016-12-20. Получено 2016-12-05.
- ^ "Stanford seeks sea urchin's secret to surviving ocean acidification | Stanford News Release". news.stanford.edu. 2013-04-08. Получено 2016-12-05.
- ^ а б Sodergren, E; Weinstock, GM; Davidson, EH; и другие. (2006-11-10). "The Genome of the Sea Urchin Strongylocentrotus purpuratus". Наука. 314 (5801): 941–952. Bibcode:2006Sci...314..941S. Дои:10.1126/science.1133609. ЧВК 3159423. PMID 17095691.
- ^ Rast, JP; Smith, LC; Loza-Coll, M; Hibino, T; Litman, GW (2006). "Genomic Insights into the Immune System of the Sea Urchin". Наука. 314 (5801): 952–6. Bibcode:2006Sci...314..952R. Дои:10.1126/science.1134301. ЧВК 3707132. PMID 17095692.
- ^ Laura Rogers-Bennett, "The Ecology of Strongylocentrotus franciscanus и Стронгилоцентротус пурпуратус" в John M. Lawrence, Edible sea urchins: biology and ecology, п. 410
- ^ а б c Davidson, Alan (2014) Оксфордский компаньон к еде. Oxford University Press, 3rd edition. С. 730–731.
- ^ John M. Lawrence, "Sea Urchin Roe Cuisine" в John M. Lawrence, Edible sea urchins: biology and ecology
- ^ "The Rise of the Sea Urchin ", Франц Лидз July 2014, Смитсоновский институт
- ^ for Puglia, Italy: Touring Club Italiano, Guida all'Italia gastronomica, 1984, с. 314; for Alexandria, Egypt: Claudia Roden, Книга о ближневосточной кухне, п. 183
- ^ Алан Дэвидсон, Средиземноморские морепродукты, п. 270
- ^ Larousse Gastronomique[страница нужна ]
- ^ Курнонский, Cuisine et vins de France, nouvelle édition, 1974, p. 248
- ^ Davidson, Alan (2014) Оксфордский компаньон к еде. Oxford University Press, 3rd edition. п. 280
- ^ Kleiman, Dena (October 3, 1990). "Scorned at Home, Maine Sea Urchin Is a Star in Japan". Нью-Йорк Таймс. п. C1.
- ^ Wassilieff, Maggy (March 2, 2009). "sea urchins". Те Ара: Энциклопедия Новой Зеландии.
- ^ Macey, Richard (November 9, 2004). "The little urchins that can command a princely price". Sydney Morning Herald.
- ^ Zatylny, Jane (6 September 2018). "Searchin' for Urchin: A Culinary Quest". Hakai magazine. Получено 10 сентября 2018.
- ^ "Sea Urchin Fishery and Overfishing", Примеры из практики TED 296, American University полный текст
- ^ Martin, R.E.; Carter, E.P.; Flick, G.J.; Davis, L.M. (2000). Marine and Freshwater Products Handbook. Тейлор и Фрэнсис. п. 268. ISBN 978-1-56676-889-4. Получено 2014-12-03.
- ^ а б Lam, Francis (2014-03-14). "California Sea Urchin Divers, Interviewed by Francis Lam". приятного аппетита. Получено 2017-03-26.
- ^ "Amulet | LIN-B37563". Схема переносных древностей. Получено 14 марта 2018.
- ^ Tullock, John H. (2008). Your First Marine Aquarium: Everything about Setting Up a Marine Aquarium, Including Conditioning, Maintenance, Selecting Fish and Invertebrates, and More. Образовательная серия Бэррона. п.63. ISBN 978-0-7641-3675-7.
- ^ McNamara, Ken (2012). "Prehistoric fossil collectors". Геологическое общество. Получено 14 марта 2018.
- ^ Marren, Peter; Mabey, Richard (2010). Ошибки Britannica. Chatto & Windus. С. 469–470. ISBN 978-0-7011-8180-2.
внешняя ссылка
- Ссылка на Всемирный регистр морских видов: Echinoidea Leske, 1778 (+ список видов)
- The sea urchin genome project
- Sea Urchin Harvesters Association – California Also, (604) 524-0322.
- The Echinoid Directory от Музей естественной истории.
- Virtual Urchin в Стэнфорде
- California Sea Urchin commission
- Introduction to the Echinoidea at UCMP Berkeley
Основной коммерческий рыболовство видовые группы | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Дикий |
|    | ||||||||||||||||
| На ферме | ||||||||||||||||||