MHC класс I - MHC class I
| Класс MHC | |
|---|---|
 Схематическое изображение MHC класса I | |
| Идентификаторы | |
| Символ | MHC класс I |
| Мембранома | 63 |
Молекулы MHC класса I являются одним из двух основных классов главный комплекс гистосовместимости (MHC) молекулы (другой MHC класс II ) и находятся на поверхность клетки из всех зародился клетки в телах позвоночные.[1][2] Они также встречаются на тромбоциты, но не на красные кровяные тельца. Их функция заключается в отображении пептидных фрагментов белков внутри клетки для цитотоксические Т-клетки; это вызовет немедленный ответ иммунной системы против определенного чужеродного антигена, отображаемого с помощью белка MHC класса I. Поскольку молекулы MHC класса I присутствуют пептиды происходит от цитозольный белки, путь презентации MHC класса I часто называют цитозольный или же эндогенный путь.[3]
У людей HLA соответствующие I классу MHC являются HLA-A, HLA-B, и HLA-C.
Функция
Молекулы MHC класса I связываются пептиды генерируется в основном из-за деградации цитозольных белков протеасома. Затем комплекс MHC I: пептид вставляется через эндоплазматический ретикулум во внешнюю плазматическую мембрану клетки. Пептид-эпитоп связывается с внеклеточными частями молекулы MHC класса I. Таким образом, функция MHC класса I заключается в отображении внутриклеточных белков для цитотоксические Т-клетки (CTL). Однако MHC класса I может также представлять пептиды, полученные из экзогенных белков, в процессе, известном как перекрестное представление.
Нормальная клетка будет отображать пептиды из нормального клеточного обмена белков на своем MHC класса I, и CTL не будут активироваться в ответ на них из-за центральных и периферических механизмов толерантности. Когда клетка экспрессирует чужеродные белки, например, после вирусной инфекции, фракция MHC класса I будет отображать эти пептиды на поверхности клетки. Следовательно, CTL, специфичные для комплекса MHC: пептид, будут распознавать и убивать представляющие клетки.
В качестве альтернативы сам по себе MHC класса I может служить в качестве ингибирующего лиганда для естественные клетки-киллеры (НК). Снижение нормальных уровней поверхностного класса I MHC, механизм, используемый некоторыми вирусами[4] и некоторые опухоли, чтобы избежать ответов CTL, активируют уничтожение NK-клеток.
ПирБ и визуальная пластичность
Парный иммуноглобулиноподобный рецептор B (PirB), рецептор, связывающий MHCI, участвует в регуляции зрительного пластичность.[5] PirB выражается в Центральная нервная система и уменьшается глазное доминирование пластичность в развитии критический период и взрослая жизнь.[5] Когда функция PirB была отменена у мутантных мышей, глазное доминирование пластичность стал более выраженным в любом возрасте.[5] У мутантных мышей с потерей функции PirB также наблюдалось повышенное пластичность после монокулярной депривации во время критический период.[5] Эти результаты предполагают, что PirB может участвовать в модуляции синаптическая пластичность в зрительная кора.
Структура
Молекулы MHC класса I представляют собой гетеродимеры, состоящие из двух полипептидных цепей, α и β.2-микроглобулин (B2M). Две цепи связаны нековалентно посредством взаимодействия B2M и α3 домен. Только α-цепь полиморфна и кодируется Ген HLA, в то время как субъединица B2M не полиморфна и кодируется Бета-2 микроглобулин ген. Α3 домен охватывает плазматическую мембрану и взаимодействует с CD8 корецептор Т-клетки. Α3-CD8 взаимодействие удерживает молекулу MHC I на месте, в то время как Рецептор Т-клеток (TCR) на поверхности цитотоксической Т-клетки связывает ее α1-α2 гетеродимерный лиганд и проверяет связанный пептид на антигенность. Α1 и α2 домены сворачиваются, образуя бороздку для связывания пептидов. Молекулы MHC класса I связывают пептиды, длина которых преимущественно составляет 8-10 аминокислот (Parham 87), но также сообщалось о связывании более длинных пептидов.[6]
Синтез
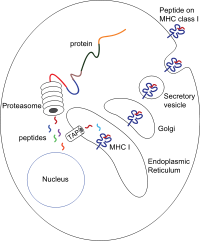
Пептиды образуются в основном в цитозоль посредством протеасома. Протеасома - это макромолекула, состоящая из 28 субъединиц, половина из которых влияет на протеолитический Мероприятия. Протеасома расщепляет внутриклеточные белки на небольшие пептиды, которые затем попадают в цитозоль. Протеасомы также могут лигировать отдельные пептидные фрагменты (называемые сплайсированными пептидами), производя последовательности, которые не являются смежными и, следовательно, не имеют линейного шаблона в геноме. Сплайсированные пептидные сегменты могут происходить из одного и того же белка (цис-сплайсинг) или из разных белков (транс-сплайсинг).[7][8] Пептиды должны быть перемещены из цитозоля в эндоплазматический ретикулум (ER), чтобы встретить молекулу MHC класса I, чей пептид-связывающий сайт находится в просвет скорой помощи. Имеют проксимальную мембрану Иг фолд
Транслокация и загрузка пептидов
Транслокация пептида из цитозоля в просвет ЭР осуществляется за счет транспортер, связанный с процессингом антигена (КРАН). ТАП является членом ABC транспортер семейство и представляет собой гетеродимерный мультимембранный полипептид, состоящий из TAP1 и TAP2. Две субъединицы образуют сайт связывания пептида и два сайта связывания АТФ, которые обращены к цитозолю. ТАП связывает пептиды на цитоплазматической стороне и перемещает их под АТФ расход в просвет ER. Затем молекула MHC класса I, в свою очередь, загружается пептидами в просвете ER.
Процесс загрузки пептида включает несколько других молекул, которые образуют большой мультимерный комплекс, называемый Комплекс загрузки пептидов[9] состоящий из TAP, тапасин, кальретикулин, калнексин, и Erp57 (PDIA3 ). Калнексин стабилизирует α-цепи MHC класса I до связывания β2m. После полной сборки молекулы MHC калнексин диссоциирует. Молекула MHC, лишенная связанного пептида, по своей природе нестабильна и требует связывания шаперонов кальретикулина и Erp57. Кроме того, тапазин связывается с молекулой MHC и служит для связывания ее с белками TAP и облегчает выбор пептида в итеративном процессе, называемом редактированием пептида.[10][11][12] таким образом облегчая увеличенную загрузку пептидов и совместную локализацию.
Как только пептид загружается в молекулу MHC класса I, комплекс диссоциирует, и он покидает ER через секреторный путь чтобы достичь поверхности клетки. Транспорт молекул MHC класса I по секреторному пути включает несколько посттрансляционные модификации молекулы MHC. Некоторые из посттрансляционных модификаций происходят в ER и включают изменения в N-гликановых областях белка, за которыми следуют обширные изменения N-гликанов в аппарат Гольджи. N-гликаны полностью созревают до того, как достигают поверхности клетки.
Удаление пептидов
Пептиды, которые не могут связывать молекулы MHC класса I в просвете эндоплазматического ретикулума (ER), удаляются из ER через сек61 канал в цитозоль,[13][14] где они могут подвергаться дальнейшему уменьшению размера и могут быть перемещены с помощью TAP обратно в ER для связывания с молекулой MHC класса I.
Например, взаимодействие sec61 с бычьим альбумин наблюдалось.[15]
Влияние вирусов
Молекулы MHC класса I загружены пептидами, образовавшимися при разложении убиквитинированный цитозольные белки в протеасомы. Поскольку вирусы индуцируют клеточную экспрессию вирусных белков, некоторые из этих продуктов маркируются для деградации, в результате чего образующиеся пептидные фрагменты попадают в эндоплазматический ретикулум и связываются с молекулами MHC I. Таким образом, MHC класса I представляет собой путь презентации антигена, когда инфицированные вирусом клетки сигнализируют Т-клеткам о том, что аномальные белки образуются в результате инфекции.
Судьба инфицированной вирусом клетки почти всегда заключается в индукции апоптоз через клеточный иммунитет, снижая риск заражения соседних клеток. В качестве эволюционного ответа на этот метод иммунного надзора многие вирусы способны подавлять или иным образом предотвращать представление молекул MHC класса I на поверхности клетки. В отличие от цитотоксических Т-лимфоцитов, естественный убийца (NK) клетки обычно инактивируются после распознавания молекул MHC I на поверхности клеток. Следовательно, в отсутствие молекул MHC I NK-клетки активируются и распознают клетку как аберрантную, что позволяет предположить, что она может быть инфицирована вирусами, пытающимися избежать иммунного разрушения. Некоторые виды рака человека также демонстрируют понижающую регуляцию MHC I, что дает трансформированным клеткам такое же преимущество выживания, как возможность избежать нормального иммунного надзора, предназначенного для уничтожения любых инфицированных или трансформированных клеток.[16]
Гены и изотипы
- Очень полиморфный
- Менее полиморфный
Эволюционная история
Гены MHC класса I произошли от самых недавний общий предок из всех челюстные позвоночные, и были обнаружены у всех живых челюстных позвоночных, изученных на данный момент.[2] С момента своего появления у челюстных позвоночных это семейство генов прошло множество различных эволюционных путей. видообразование события произошли. Однако есть задокументированные случаи трансвидового полиморфизмы в генах MHC класса I, где конкретный аллель в эволюционно родственном гене MHC класса I остается у двух видов, вероятно, из-за сильного опосредованного патогенами балансирующий выбор к патогены которые могут заразить оба вида.[17] Рождение и смерть Эволюция - одно из механистических объяснений размера семейства генов MHC класса I.
Рождение и смерть генов MHC класса I
Эволюция рождения и смерти утверждает, что дупликация гена События заставляют геном содержать несколько копий гена, которые затем могут претерпевать отдельные эволюционные процессы. Иногда эти процессы приводят к псевдогенизация (смерть) одной копии гена, хотя иногда этот процесс приводит к появлению двух новых генов с дивергентной функцией.[18] Вероятно, что локусы MHC класса Ib человека (HLA-E, -F и -G), а также псевдогены MHC класса I произошли от локусов MHC класса Ia (HLA-A, -B и -C) в этом рождении. а-смертельный процесс.[19]
Рекомендации
- ^ Hewitt EW (октябрь 2003 г.). «Путь презентации антигена MHC класса I: стратегии уклонения от вирусного иммунитета». Иммунология. 110 (2): 163–9. Дои:10.1046 / j.1365-2567.2003.01738.x. ЧВК 1783040. PMID 14511229.
- ^ а б Кульски Дж. К., Шиина Т., Анзай Т., Кохара С., Иноко Х. (декабрь 2002 г.). «Сравнительный геномный анализ MHC: эволюция блоков дупликации I класса, разнообразие и сложность от акулы к человеку». Иммунологические обзоры. 190: 95–122. Дои:10.1034 / j.1600-065x.2002.19008.x. PMID 12493009.
- ^ http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/H/HLA.html#Class_I_Histocompatibility_Molecules Страницы биологии Кимбалла, молекулы гистосовместимости
- ^ Хансен TH, Бувье М (июль 2009 г.). «Презентация антигена класса I MHC: обучение стратегиям уклонения от вирусов». Обзоры природы. Иммунология. 9 (7): 503–13. Дои:10.1038 / nri2575. PMID 19498380.
- ^ а б c d Syken J, Grandpre T., Kanold PO, Shatz CJ (сентябрь 2006 г.). «PirB ограничивает пластичность глазного доминирования в зрительной коре». Наука. 313 (5794): 1795–800. Bibcode:2006Научный ... 313.1795S. Дои:10.1126 / science.1128232. PMID 16917027.
- ^ Берроуз С.Р., Россджон Дж., Маккласки Дж. (Январь 2006 г.). «Неужели мы слишком ограничились в картировании эпитопов CTL?». Тенденции в иммунологии. 27 (1): 11–6. Дои:10.1016 / j.it.2005.11.001. PMID 16297661.
- ^ Фариди, Пуйя; Ли, Чен; Ramarathinam, Sri H .; Вивиан, Джулиан П .; Иллинг, Патрисия Т .; Mifsud, Nicole A .; Айяла, Рошель; Сун, Цзяннин; Gearing, Linden J .; Герцог, Пол Дж .; Тернетт, Никола; Россджон, Джейми; Крофт, Натан П .; Перселл, Энтони В. (12 октября 2018 г.). «Подмножество пептидов HLA-I не является геномным шаблоном: данные о цис- и транс-сплайсированных пептидных лигандах» (PDF). Научная иммунология. 3 (28): eaar3947. Дои:10.1126 / sciimmunol.aar3947. PMID 30315122.
- ^ Лиепе, Джулиана; Марино, Фабио; Сидней, Джон; Джеко, Анита; Бантинг, Дэниел Э .; Сетте, Алессандро; Kloetzel, Peter M .; Штумпф, Майкл П. Х .; Heck, Albert J. R .; Мишто, Микеле (21 октября 2016 г.). «Большая часть лигандов HLA класса I представляет собой сплайсированные пептиды, полученные с помощью протеасом» (PDF). Наука. 354 (6310): 354–358. Bibcode:2016Научный ... 354..354L. Дои:10.1126 / science.aaf4384. HDL:10044/1/42330. PMID 27846572.
- ^ Blees A, Januliene D, Hofmann T., Koller N, Schmidt C, Trowitzsch S, Moeller A, Tampé R (ноябрь 2017 г.). «Структура комплекса загрузки пептида MHC-I человека». Природа. 551 (7681): 525–528. Bibcode:2017Натура.551..525Б. Дои:10.1038 / природа24627. PMID 29107940.
- ^ Ховарт М., Уильямс А., Толструп А.Б., Эллиотт Т. (август 2004 г.). «Тапазин усиливает представление пептида MHC класса I в соответствии с периодом полужизни пептида». Труды Национальной академии наук Соединенных Штатов Америки. 101 (32): 11737–42. Bibcode:2004PNAS..10111737H. Дои:10.1073 / pnas.0306294101. ЧВК 511045. PMID 15286279.
- ^ Вирш П.А., Крессвелл П. (август 2007 г.). «Селективная загрузка высокоаффинных пептидов на молекулы класса I главного комплекса гистосовместимости гетеродимером тапазин-ERp57». Иммунология природы. 8 (8): 873–81. Дои:10.1038 / ni1485. PMID 17603487.
- ^ Thirdborough SM, Роддик Дж. С., Рэдклифф Дж. Н., Ховарт М., Стивенсон Ф. К., Эллиот Т. (февраль 2008 г.). «Тапазин формирует иерархию иммунодоминантности в соответствии с кинетической стабильностью комплексов пептид-MHC класса I». Европейский журнал иммунологии. 38 (2): 364–9. Дои:10.1002 / eji.200737832. PMID 18196518.
- ^ Купманн Дж., Альбринг Дж., Хютер Э, Бульбук Н., Шпее П., Нифьес Дж., Хэммерлинг Дж. Дж., Момбург Ф. и др. (Июль 2000 г.). «Экспорт антигенных пептидов из эндоплазматического ретикулума пересекается с ретроградной транслокацией белка через канал Sec61p». Иммунитет. 13 (1): 117–27. Дои:10.1016 / S1074-7613 (00) 00013-3. PMID 10933400.
- ^ Альбринг Дж., Купманн Дж. О., Хэммерлинг Дж. Дж., Момбург Ф. (январь 2004 г.). «Ретротранслокация тяжелой цепи MHC класса I из эндоплазматического ретикулума в цитозоль зависит от поступления АТФ в просвет ER». Молекулярная иммунология. 40 (10): 733–41. Дои:10.1016 / j.molimm.2003.08.008. PMID 14644099.
- ^ Имаи Дж., Хасэгава Х., Маруя М., Коясу С., Яхара И. (январь 2005 г.). «Экзогенные антигены процессируются посредством деградации, связанной с эндоплазматическим ретикулумом (ERAD), при перекрестной презентации дендритными клетками». Международная иммунология. 17 (1): 45–53. Дои:10.1093 / intimm / dxh184. PMID 15546887.
- ^ Ван З, Чжан Л., Цяо А., Ватсон К., Чжан Дж., Фань Г. Х. (февраль 2008 г.). «Активация CXCR4 запускает убиквитинирование и подавление основного комплекса гистосовместимости класса I (MHC-I) на клетках эпителиоидной карциномы HeLa». Журнал биологической химии. 283 (7): 3951–9. Дои:10.1074 / jbc.m706848200. PMID 18083706.
- ^ Азеведо Л., Серрано С., Аморим А., Купер Д. Н. (сентябрь 2015 г.). «Межвидовой полиморфизм у людей и человекообразных обезьян обычно поддерживается за счет сбалансированного отбора, который модулирует иммунный ответ хозяина». Геномика человека. 9: 21. Дои:10.1186 / s40246-015-0043-1. ЧВК 4559023. PMID 26337052.
- ^ Ней М., Руни А.П. (14 ноября 2005 г.). «Согласованная эволюция мультигенных семей и рождение-смерть». Ежегодный обзор генетики. 39 (1): 121–52. Дои:10.1146 / annurev.genet.39.073003.112240. ЧВК 1464479. PMID 16285855.
- ^ Хьюз А.Л. (март 1995 г.). «Происхождение и эволюция псевдогенов HLA класса I». Молекулярная биология и эволюция. 12 (2): 247–58. Дои:10.1093 / oxfordjournals.molbev.a040201. PMID 7700152.
внешняя ссылка
- Гистосовместимость + Антигены + Класс + I в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- MHC + Класс + I + Гены в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)