Пептидогликан - Peptidoglycan
Пептидогликан или же Murein это полимер состоящий из сахара и аминокислоты который образует сетчатый слой за пределами плазматическая мембрана из большинства бактерии, формируя клеточная стенка. Сахарный компонент состоит из чередующихся остатков β- (1,4) связанных N-ацетилглюкозамин (NAG) и N-ацетилмурамовая кислота (ДН). Прикреплен к N-ацетилмурамовая кислота представляет собой пептидную цепь из трех-пяти аминокислот. Пептидная цепь может быть поперечно связана с пептидной цепью другой цепи, образующей трехмерный сетчатый слой.[1] Пептидогликан выполняет структурную роль в стенке бактериальной клетки, придавая структурную прочность, а также противодействуя осмотическое давление из цитоплазма. Пептидогликан также участвует в двойное деление во время размножения бактериальных клеток.
Слой пептидогликана значительно толще в Грамположительный бактерии (От 20 до 80 нанометров), чем в Грамотрицательный бактерии (от 7 до 8 нанометров).[2] В зависимости от условий роста pH пептидогликан образует от 40 до 90% клеточная стенка с сухой вес грамположительных бактерий, но только около 10% грамотрицательных штаммов. Таким образом, наличие высоких уровней пептидогликана является основным фактором, определяющим характеристику бактерий как грамположительных.[3] У грамположительных штаммов это важно в ролях привязанности и серотипирование целей.[4] Как для грамположительных, так и для грамотрицательных бактерий частицы размером примерно 2 нм могут проходить через пептидогликан.[5]
Структура
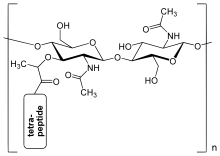
Слой пептидогликана в стенке бактериальной клетки представляет собой кристаллическая решетка структура образована линейными цепями двух чередующихся амино сахара, а именно N-ацетилглюкозамин (GlcNAc или NAGA) и N-ацетилмурамовая кислота (MurNAc или NAMA). Чередующиеся сахара связаны β- (1,4) -гликозидная связь. Каждый MurNAc прикреплен к короткому (от 4 до 5 остатков) аминокислота цепь, содержащая L-аланин, D-глютаминовая кислота, мезо-диаминопимелиновая кислота, и D-аланин в случае кишечная палочка (грамотрицательная бактерия) или L-аланин, D-глютамин, L-лизин, и D-аланин с 5-глицин межмостик между тетрапептидами в случае Золотистый стафилококк (грамположительная бактерия). Пептидогликан - один из важнейших источников D-аминокислоты в природе.
Сшивание между аминокислоты в различных линейных аминокислотных цепях сахара происходит с помощью фермента DD-транспептидаза В результате получается прочная и жесткая трехмерная структура. Конкретная аминокислотная последовательность и молекулярная структура меняются в зависимости от бактериального разновидность.[6]

Строение пептидогликана. НАГ = N-ацетилглюкозамин (также называемый GlcNAc или NAGA), NAM = N-ацетилмурамовая кислота (также называемый MurNAc или NAMA).

Пенициллин-связывающий белок формирование поперечных связей в вновь образованной клеточной стенке бактерий.


Биосинтез
Мономеры пептидогликана синтезируются в цитозоль и затем прикрепляются к мембранному носителю бактопренол. Бактопренол переносит мономеры пептидогликана через клеточную мембрану, где они вставляются в существующий пептидогликан.[7]
На первом этапе синтеза пептидогликана глутамин, который является аминокислотой, отдает аминогруппу сахару, фруктозо-6-фосфат. Это превращает фруктозо-6-фосфат в глюкозамин-6-фосфат. На втором этапе ацетильная группа переносится из ацетил-КоА к аминогруппе глюкозамин-6-фосфата, создавая N-ацетил-глюкозамин-6-фосфат.[8] На третьем этапе процесса синтеза N-ацетил-глюкозамин-6-фосфат изомеризуется, что изменит N-ацетилглюкозамин-6-фосфат до N-ацетил-глюкозамин-1-фосфат.[8]
На шаге 4 N-ацетил-глюкозамин-1-фосфат, который теперь является монофосфатом, атакует UTP. Уридинтрифосфат, который является пиримидин нуклеотид, обладает способностью действовать как источник энергии. В этой конкретной реакции после того, как монофосфат атакует UTP, выделяется неорганический пирофосфат и заменяется монофосфатом, образуя UDP-N-ацетилглюкозамин (2,4). (Когда UDP используется в качестве источника энергии, он выделяет неорганический фосфат.) Эта начальная стадия используется для создания предшественника NAG в пептидогликане.
На этапе 5 часть UDP-N-ацетилглюкозамина (UDP-GlcNAc) превращается в UDP-MurNAc (UDP-N-ацетилмурамовая кислота) путем добавления лактильной группы к глюкозамину. Также в этой реакции гидроксильная группа C3 удаляет фосфат из альфа-углерода фосфоенолпируват. Это создает так называемое производное енола, которое будет восстановлено до «лактильного фрагмента» под действием НАДФН на шестой стадии.[8]
На этапе 7 UDP-MurNAc превращается в пентапептид UDP-MurNAc путем добавления пяти аминокислот, обычно включая дипептид. D-аланил-D-аланин.[8] Каждая из этих реакций требует источника энергии АТФ.[8] Все это называется этапом один.
Вторая стадия происходит в цитоплазматической мембране. Именно в мембране липидный носитель, называемый бактопренолом, переносит предшественники пептидогликана через клеточную мембрану. Бактопренол будет атаковать UDP-MurNAc penta, создавая PP-MurNac penta, которая теперь является липидом. Затем UDP-GlcNAc транспортируется в MurNAc, создавая Lipid-PP-MurNAc пента-GlcNAc, дисахарид, также являющийся предшественником пептидогликана.[8] Как эта молекула транспортируется через мембрану, до сих пор не понятно. Однако, как только он там оказывается, он добавляется к растущей цепи гликана.[8] Следующая реакция известна как трангликозилирование. В этой реакции гидроксильная группа GlcNAc будет присоединяться к MurNAc в гликане, что вытесняет липид-PP из гликановой цепи. За это отвечает фермент трансгликозилаза.[8]
Торможение
Немного антибактериальные препараты Такие как пенициллин мешают выработке пептидогликана путем связывания с бактериальными ферментами, известными как пенициллин-связывающие белки или же DD-транспептидазы.[4] Пенициллин-связывающие белки образуют связи между поперечными связями олигопептидов в пептидогликане. Для размножения бактериальной клетки через двойное деление, более миллиона субъединиц пептидогликана (NAM-NAG + олигопептид) должны быть присоединены к существующим субъединицам.[9] Мутации в генах, кодирующих транспептидазы, которые приводят к снижению взаимодействий с антибиотиком, являются важным источником новых устойчивость к антибиотикам.[10]Мурамицины представляют собой подкласс нуклеозидных антибиотиков, которые действуют как конкурентные ингибиторы природного субстрата UDP-MurNAc-пентапептида (UM5A) фосфо-N-ацетилмурамоил-пентапептида транслоказы (MraY).[11]
Лизоцим, который содержится в слезах и составляет часть тела врожденная иммунная система оказывает антибактериальный эффект, разрывая β- (1,4) -гликозидные связи в пептидогликане (см. выше).
Сходство с псевдопептидогликаном
Немного археи иметь аналогичный слой псевдопептидогликан (также известный как псевдомуреин), в котором сахарные остатки связаны β- (1,3) N-ацетилглюкозамин и N-ацетилталозаминуроновая кислота. Это делает клеточные стенки таких архей нечувствительными к лизоцим.[12]
Рекомендации
- ^ Анимация синтеза слоя пептидогликана
- ^ Перселл А. (18 марта 2016 г.). «Бактерии». Основы биологии.
- ^ К. Майкл Хоган. 2010 г. Бактерии. Энциклопедия Земли. ред. Сидни Драгган и Си Джей Кливленд, Национальный совет по науке и окружающей среде, Вашингтон, округ Колумбия
- ^ а б Солтон MR, Ким KS (1996). "Структура". В Baron S et al. (ред.). Структура. В: Медицинская микробиология Барона (4-е изд.). Univ Техасского медицинского отделения. ISBN 978-0-9631172-1-2.
- ^ Демчик PH, Кох А.Л. (1 февраля 1996 г.). «Проницаемость стеночной ткани для кишечной палочки и Bacillus subtilis». Журнал бактериологии. 178 (3): 768–73. Дои:10.1128 / jb.178.3.768-773.1996. ЧВК 177723. PMID 8550511.
- ^ Райан К.Дж., Рэй К.Г., ред. (2004). Шеррис Медицинская микробиология (4-е изд.). Макгроу Хилл. ISBN 978-0-8385-8529-0.
- ^ «II. ПРОКАРИОТИЧЕСКАЯ КЛЕТКА: БАКТЕРИИ». Архивировано из оригинал 26 июля 2010 г.. Получено 1 мая 2011.
- ^ а б c d е ж грамм час Уайт, Д. (2007). Физиология и биохимия прокариот (3-е изд.). Нью-Йорк: Oxford University Press Inc.
- ^ Бауман Р. (2007). 2-е (изд.). Микробиология с болезнями по таксономии. Бенджамин Каммингс. ISBN 978-0-8053-7679-1.
- ^ Spratt BG (апрель 1994 г.). «Устойчивость к антибиотикам, опосредованная изменением мишеней». Наука. 264 (5157): 388–93. Дои:10.1126 / science.8153626. PMID 8153626. S2CID 30578841.
- ^ Саид Малек Заде (2 сентября 2020 г.). «Теоретическое исследование межмолекулярных взаимодействий между критическими остатками мембранного белка MraYAA и перспективным антибиотиком мурамицином D2». СКУД Омега. 5 (36): 22739–22749. Дои:10.1021 / acsomega.0c01551.
- ^ Мэдиган М. Т., Дж. М. Мартинко, П. В. Данлэп и Д. П. Кларк. Брок-биология микроорганизмов. 12-е изд. Сан-Франциско, Калифорния: Пирсон / Бенджамин Каммингс, 2009.
внешняя ссылка
- Схематическое изображение структур пептидогликана.
- Структура MurNAc 6-фосфатгидролазы (MurQ) из Haemophilus influenzae со связанным ингибитором.
| Медицинское микробиология | |||||||
|---|---|---|---|---|---|---|---|
| Биохимия и экология |
| ||||||
| Форма | |||||||
| Структура |
| ||||||
| Таксономия и эволюция | |||||||