Топоизомераза типа II - Type II topoisomerase
| ДНК-топоизомераза II (АТФ-гидролизующий) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Структура 42 кДа фрагмента N-концевой АТФазы и трансдукторных доменов ДНК-гиразы, гомологичной всем другим топоизомеразам типа IIA. | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 5.6.2.2 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
Топоизомеразы типа II находятся топоизомеразы которые разрезают обе нити спирали ДНК одновременно, чтобы управлять ДНК путаницы и суперспирали. Они используют гидролиз АТФ, В отличие от Топоизомераза I типа. В этом процессе эти ферменты изменяют номер ссылки кольцевой ДНК на ± 2.
Функция
После разрезания концы ДНК разделяются, и второй дуплекс ДНК пропускается через разрыв. После пассажа разрезанная ДНК повторно лигируется. Эта реакция позволяет топоизомеразам типа II увеличивать или уменьшать связующее число петли ДНК на 2 единицы и способствует распутыванию хромосом. Для реакций увеличения суперспирализации требуются две молекулы АТФ. Например, ДНК-гираза, топоизомераза типа II, наблюдаемая в Кишечная палочка и большинство других прокариоты, вводит отрицательные суперспирали и уменьшает число зацеплений на 2. Gyrase также может удалять узлы с бактериальная хромосома. Наряду с гиразой большинство прокариот также содержат топоизомеразу второго типа IIA, называемую топоизомеразой IV. Гираза и топоизомераза IV различаются своими С-концевыми доменами, что, как полагают, определяет субстратную специфичность и функциональность этих двух ферментов. Футпринтинг указывает на то, что гираза, которая образует след из 140 пар оснований и обертывает ДНК, вносит отрицательные суперспирали, в то время как топоизомераза IV, которая формирует след из 28 пар оснований, не обертывает ДНК.
Топоизомераза эукариот типа II не может создавать суперспирали; это может только расслабить их.
Роль топоизомераз типа IIB менее изучена. В отличие от топоизомераз типа IIA, топоизомеразы типа IIB не могут упростить топологию ДНК (см. Ниже), но они имеют несколько общих структурных особенностей с топоизомеразами типа IIA.
Упрощение топологии
Топоизомеразы типа IIA важны для разделения запутанных дочерних цепей во время репликации. Считается, что эту функцию выполняет топоизомераза II у эукариот и топоизомераза IV у прокариот. Неспособность разделить эти нити приводит к гибели клеток. Топоизомеразы типа IIA обладают особой способностью релаксировать ДНК до состояния ниже термодинамического равновесия, в отличие от топоизомераз типа IA, IB и IIB. Эта способность, известная как упрощение топологии, была впервые обнаружена Рыбенковым и др.[1] Гидролиз АТФ способствует этому упрощению, но четкого молекулярного механизма для этого упрощения все еще нет. Было предложено несколько моделей для объяснения этого явления, в том числе две модели, которые полагаются на способность топоизомераз типа IIA распознавать изогнутые дуплексы ДНК.[2] Биохимия, электронная микроскопия и недавние структуры топоизомеразы II, связанной с ДНК, показывают, что топоизомеразы типа IIA связываются на вершинах ДНК, что подтверждает эту модель.
Классификация
Существует два подкласса топоизомераз типа II, типа IIA и IIB.
- Топоизомеразы типа IIA включают ферменты ДНК-гираза, эукариотическая топоизомераза II (topo II) и бактериальная топоизомераза IV (топо IV). Эти ферменты охватывают все сферы жизни и необходимы для функционирования.[3]
- Топоизомеразы типа IIB структурно и биохимически различаются и включают единственного члена семейства, топоизомеразу VI (topo VI). Топоизомеразы типа IIB обнаружены у архей и некоторых высших растений.
У некоторых организмов есть две изоформы топоизомеразы II: альфа и бета. В раки, топоизомераза II-альфа высоко экспрессируется в сильно пролиферирующих клетках. При некоторых видах рака, таких как опухоли оболочек периферических нервов, высокая экспрессия кодируемого им белка также связана с плохой выживаемостью пациентов.
Два класса топоизомераз обладают схожим механизмом прохождения цепи и доменной структурой (см. Ниже), однако у них также есть несколько важных различий. Топоизомеразы типа IIA образуют двухцепочечные разрывы с выступами из четырех пар оснований, тогда как топоизомеразы типа IIB образуют двухцепочечные разрывы с двумя выступами оснований.[4] Кроме того, топоизомеразы типа IIA способны упростить топологию ДНК,[1] в то время как топоизомеразы типа IIB - нет.[5]
Структура
Тип IIA

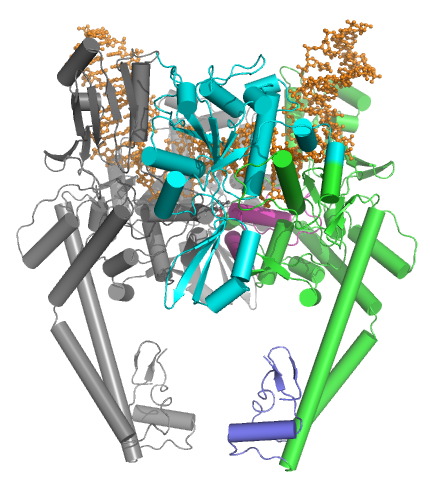
Топоизомеразы типа IIA состоят из нескольких ключевых мотивов:
- N-концевой GHKL АТФаза домен (для гиразы, Hsp, киназы и MutL),
- а Домен Toprim (а Россманн фолд подкласс), который существует как в топоизомеразах типа II, так и в топоизомеразах типа IA и бактериальной примазе (DnaG),
- центральное ДНК-связывающее ядро (которое структурно образует структуру в форме сердца), и
- вариабельный С-концевой домен.
Топоизомеразы эукариот II типа являются гомодимерами (A2), а прокариотические топоизомеразы II типа - гетеротетрамеры (A2B2). Прокариоты имеют домен АТФазы и складку Топрим на одном полипептиде (Pfam PF00204 ), в то время как ядро расщепления ДНК и CTD лежит на втором полипептиде (Pfam PF00521 ). Для гиразы первый полипептид называется GyrB, а второй полипептид называется GyrA. Для topo IV первый полипептид называется ParE, а второй полипептид - ParC. Обе сигнатуры Pfam обнаружены в одноцепочечной эукайотической топоизомеразе.
Структуры N-концевого АТФазного домена гиразы[6] и дрожжевая топоизомераза II[7] были решены в комплексе с AMPPNP (аналог АТФ), показывая, что два домена АТФазы димеризуются с образованием замкнутой конформации. Для гираза в структуре есть значительное отверстие в середине, которое, как предполагается, предназначено для размещения Т-образного сегмента.
Связывание домена АТФазы со складкой Toprim представляет собой спиральный элемент, известный как домен-преобразователь. Считается, что этот домен сообщает о нуклеотидном состоянии домена АТФазы остальной части белка. Модификации этого домена влияют на активность топоизомеразы, а структурная работа, проделанная группой Verdine, показывает, что состояние АТФ влияет на ориентацию домена-преобразователя.[8]
Центральное ядро белка содержит складку Toprim и ДНК-связывающее ядро, которое содержит домен крылатой спирали (WHD), часто называемый доменом CAP, поскольку он был впервые идентифицирован как похожий на WHD белка-активатора катаболита. Каталитический тирозин находится на этом WHD. Фолда Топрим - это складка Россмана, которая содержит три инвариантных кислотных остатка, которые координируют ионы магния, участвующие в расщеплении ДНК и лигировании ДНК.[9] Строение складки Toprim и ДНК-связывающего ядра дрожжевой топоизомеразы II было впервые решено Бергером и Вангом,[10] и первое ДНК-связывающее ядро гиразы было решено Morais Cabral et al.[11] Структура, решенная Бергером, позволила получить важные сведения о функции фермента. ДНК-связывающее ядро состоит из WHD, которая ведет к башенному домену. Область спиральной катушки ведет к С-концевому домену, который формирует главный интерфейс димера для этого кристаллического состояния (часто называемого С-затвором). В то время как исходная структура топоизомеразы II показывает ситуацию, когда WHD разделены большим расстоянием, структура гиразы показывает закрытую конформацию, где WHD близки.
Ядро топоизомеразы II было позже решено в новых конформациях, в том числе Fass et al.[12] и один Донг и др.[13] Структура Fass показывает, что домен Toprim является гибким и что эта гибкость может позволить домену Toprim координироваться с WHD с образованием компетентного комплекса расщепления. В конечном итоге это было подтверждено Dong et al. структура, которая была решена в присутствии ДНК. Эта последняя структура показала, что домен Toprim и WHD образуют комплекс расщепления, очень похожий на комплекс топоизомераз типа IA, и указали, как ДНК-связывание и расщепление могут быть разъединены, а структура показала, что ДНК изогнута на ~ 150 градусов через инвариантный изолейцин (в топоизомеразе II это I833, а в гиразе I172). Этот механизм изгиба очень похож на механизм интеграции фактора хозяина (IHF) и HU, двух архитектурных белков у бактерий. Кроме того, в то время как предыдущие структуры ДНК-связывающего ядра имели закрытые ворота C, эта структура захватила ворота открытыми, что является ключевым этапом в механизме двух ворот (см. Ниже).
Совсем недавно было решено несколько структур связанных с ДНК структур в попытке понять как химический механизм расщепления ДНК, так и структурную основу ингибирования топоизомеразы антибактериальными ядами. Первая полная архитектура Кишечная палочка ДНК-гираза была обнаружена с помощью криоэлектронной микроскопии с разрешением, близким к атомному.[14] Нуклеопротеиновый комплекс был захвачен длинным дуплексом ДНК и гепотидацином, новым ингибитором бактериальной топоизомеразы.
С-концевая область прокариотических топоизомераз была определена для многих видов. Первая структура C-концевого домена гиразы была решена Corbett et al.[15] и C-концевой домен топоизомеразы IV был решен Corbett et al.[5] Эти структуры сформировали новый бета-ствол, который изгибает ДНК, оборачивая вокруг себя нуклеиновую кислоту. Изгиб ДНК под действием гиразы был предложен в качестве ключевого механизма способности гиразы вводить в ДНК отрицательные суперспирали. Это согласуется с данными следа, которые показывают, что гираза имеет след 140 пар оснований. CTD как гиразы, так и топоизомеразы IV изгибают ДНК, но только гираза вводит отрицательные суперспирали.
В отличие от функции C-концевого домена прокариотических топоизомераз, функция C-концевого участка эукариотической топоизомеразы II до сих пор не ясна. Исследования показали, что эта область регулируется фосфорилированием, и это модулирует активность топоизомеразы, однако для изучения этого необходимы дополнительные исследования.
Тип IIB
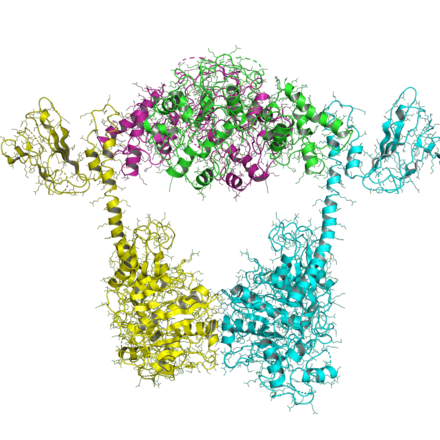
Организация топоизомераз типа IIB аналогична организации типа IIA, за исключением того, что все типы IIB имеют два гена и образуют гетеротетрамеры. Один ген, названный topo VI-B (так как он похож на gyrB), содержит домен АТФазы, домен-преобразователь (Pfam PF09239 ), и C-концевой Ig-фолд-подобный H2TH домен (Pfam PF18000 ). Второй ген, названный topo VI-A (Pfam PF04406 ), содержит WHD и домен Toprim.
АТФазный домен topo VI B был расшифрован в множественных нуклеотидных состояниях.[16] Он очень похож на домен GHKL topo II и MutL и показывает, что состояние нуклеотида (АДФ по сравнению с АТФ) влияет на ориентацию домена-преобразователя (и 1MX0).
Структура топо VI-A была решена Bergerat et al.[17] показывая, что складки HTH и Toprim имели новую конформацию по сравнению с конформацией topo IIA.
Недавно была решена структура комплекса topo VI A / B, показывающая открытую и закрытую конформации, два состояния, которые предсказываются в механизме двух ворот (см. Ниже). Эти структуры, одна из которых представляет собой рентгеновскую кристаллическую структуру, а другая - реконструкцию малоуглового рентгеновского рассеяния (SAXS), показывают, что домен АТФазы может быть как открытым, так и закрытым.[18]
Механизм действия
Прямой переход
Топоизомераза типа IIA действует через механизм «двух ворот» (хотя это историческое обозначение), механизм, поддерживаемый биохимией.[19] а также структурными работами.[20]
Нить ДНК, называемая воротами, или G-сегментами, связана с центральными воротами связывания ДНК (ДНК-воротами). Вторая цепь ДНК, называемая транспортной, или Т-сегментом, захватывается димеризацией N-концевого домена АТФазы (ворота АТФазы), когда две молекулы АТФ связываются. Гидролиз АТФ и высвобождение неорганического фосфата приводит к расщеплению G-сегмента, поскольку каталитические тирозины образуют ковалентную фосфотирозиновую связь с 5'-концом ДНК. Это создает выступ из четырех оснований и двухцепочечный разрыв в G-сегменте. Когда ворота связывания ДНК отделяются, Т-сегмент переносится через G-сегмент. G-сегмент герметичен, что приводит к открытию ворот C-терминала (или C-образного элемента), что позволяет освободить T-сегмент. Выпуск ADP продукта приводит к перезагрузке системы и позволяет захватить второй T-сегмент.
Топоизомеразы типа IIB действуют аналогичным образом, за исключением того, что белок образует выступ из двух оснований в G-сегменте, а С-концевой гейт полностью отсутствует.
Расщепление ДНК
В механизме пассажа цепочки расщепление ДНК является ключевым фактором, позволяющим Т-сегменту проходить через G-сегмент. Механизм расщепления ДНК топоизомеразами типа IIA недавно был в центре внимания многих биохимических и структурно-биологических исследований.
Цепочка
Цепочка это процесс, с помощью которого две кольцевые нити ДНК соединяются вместе как звенья цепи. Это происходит после репликации ДНК, когда две одиночные нити сцеплены и все еще могут реплицироваться, но не могут разделиться на две дочерние клетки. Поскольку топоизомеры типа II разрывают двойную цепь, они могут исправить это состояние (топоизомеразы типа I могут сделать это, только если уже имеется одноцепочечный разрыв), и правильное число хромосом может остаться в дочерних клетках. Линейная ДНК в эукариоты так долго, что их можно рассматривать как бесконечные; топоизомеразы типа II необходимы по той же причине.
Торможение
Небольшие молекулы, нацеленные на топоизомеразу типа II, делятся на два класса: ингибиторы и яды. Из-за их частого присутствия в пролиферирующих эукариотических клетках ингибиторы топоизомераз типа II широко изучаются и используются в качестве противораковых препаратов.[21]
- Ингибиторы топоизомеразы типа II включают: HU-331, ICRF-187, ICRF-193, и митиндомид. Эти молекулы действуют путем ингибирования активности АТФазы, действуя как неконкурентные ингибиторы АТФ. Это было показано структурными исследованиями.[7] и биохимические исследования, проведенные группой Линдсли.
- Яды топоизомераз типа II включают: этопозид, новобиоцин, хинолоны (включая ципрофлоксацин ), и тенипозид. Эти небольшие молекулы нацелены на комплекс ДНК-белок. Некоторые из этих молекул приводят к усиленному расщеплению, тогда как другие, такие как этопозид, ингибируют религирование.
Экспериментальный противоопухолевый препарат м-AMSA (4 '- (9'-акридиниламино) метансульфон-м-анизидид) также ингибирует топоизомеразу 2 типа.[22]
Яды топоизомеразы широко используются как в противораковой, так и в антибактериальной терапии. Хотя антибактериальные соединения, такие как ципрофлоксацин, нацелены на бактериальную гиразу, они не могут ингибировать эукариотический топоизомеразы типа IIA. Кроме того, устойчивые к лекарствам бактерии часто имеют точечную мутацию в гираза (Serine79Alanine в E. coli), который делает хинолоны неэффективными.[нужна цитата ] Недавние структурные исследования привели к открытию соединения, которое больше не зависит от этого остатка и, следовательно, имеет эффективность против устойчивых к лекарствам бактерий.[нужна цитата ]
Бактериофаг Т4 гираза
Гираза бактериофага (фага) Т4 (топоизмераза типа II) представляет собой мультисубъединичный белок, состоящий из продуктов генов 39, 52 и, вероятно, 60.[23][24] Он катализирует расслабление отрицательно или положительно сверхспиральной ДНК и используется в фаге. Репликация ДНК во время заражения Кишечная палочка бактериальный хозяин.[25] Белок гена 52 фага гомологичен белку Кишечная палочка субъединица gyrase gyrA[26] и белок гена фага 39 имеет гомологию с субъединицей gyr B.[27] Поскольку хозяин Кишечная палочка ДНК-гираза может частично компенсировать потерю продуктов гена фага Т4, мутанты, дефектные по генам 39, 52 или 60, не полностью отменяют репликацию фаговой ДНК, а скорее задерживают ее начало.[25] Скорость удлинения ДНК при таких мутантных инфекциях не ниже, чем у дикого типа.[28] Мутанты, дефектные по генам 39, 52 или 60, показывают повышенное генетическая рекомбинация а также увеличение количества замен и делеций оснований мутация предполагая, что синтез ДНК с компенсацией хозяина менее точен, чем управляемый фагом дикого типа.[29] Мутант, дефектный по гену 39, проявляет повышенную чувствительность к инактивации ультрафиолетовый облучение на стадии фаговой инфекции после инициации репликации ДНК при множественных копиях фага хромосома присутствуют.[30] Мутанты, дефектные по генам 39, 52 и 60, обладают пониженной способностью выполнять реактивацию множественности, форму рекомбинационной репарации, которая может иметь дело с различными типами повреждений ДНК.[31] Гираза, заданная геномом неинфицированных Кишечная палочка также, по-видимому, участвует в рекомбинационной репарации, обеспечивая точку инициации для обмена реципрокной цепи, управляемого белком RecA.[32]
Рекомендации
- ^ а б Рыбенков В.В., Ульспергер С., Вологодский А.В., Коццарелли Н.Р. (август 1997 г.). «Упрощение топологии ДНК ниже равновесных значений за счет топоизомераз типа II». Наука. Нью-Йорк, штат Нью-Йорк. 277 (5326): 690–3. Дои:10.1126 / science.277.5326.690. PMID 9235892.
- ^ Вологодский А.В., Чжан В., Рыбенков В.В., Подтележников А.А., Субраманиан Д., Гриффит Д.Д., Коццарелли Н.Р. (март 2001 г.). «Механизм упрощения топологии ДНК-топоизомеразами II типа». Труды Национальной академии наук Соединенных Штатов Америки. 98 (6): 3045–9. Bibcode:2001PNAS ... 98.3045V. Дои:10.1073 / pnas.061029098. ЧВК 30604. PMID 11248029.
- ^ Рис Р.Дж., Максвелл А. (январь 1991 г.). «ДНК-гираза: структура и функции». Критические обзоры в биохимии и молекулярной биологии. 26 (3–4): 335–75. Дои:10.3109/10409239109114072. PMID 1657531.
- ^ Бюлер C, Леббинк JH, Bocs C, Ladenstein R, Forterre P (октябрь 2001 г.). «ДНК-топоизомераза VI генерирует АТФ-зависимые двухцепочечные разрывы с двухнуклеотидными выступами». Журнал биологической химии. 276 (40): 37215–22. Дои:10.1074 / jbc.M101823200. PMID 11485995. S2CID 24354635.
- ^ а б PDB: 1звт; Корбетт К.Д., Шеффлер А.Д., Томсен Н.Д., Бергер Дж. М. (август 2005 г.). «Структурные основы субстратной специфичности ДНК-топоизомеразы IV». Журнал молекулярной биологии. 351 (3): 545–61. Дои:10.1016 / j.jmb.2005.06.029. PMID 16023670.
- ^ Вигли Д.Б., Дэвис Г.Дж., Додсон Э.Дж., Максвелл А., Додсон Г. (июнь 1991 г.). «Кристаллическая структура N-концевого фрагмента белка ДНК-гиразы B». Природа. 351 (6328): 624–9. Bibcode:1991 Натур.351..624Вт. Дои:10.1038 / 351624a0. PMID 1646964. S2CID 4373125.
- ^ а б PDB: 1ПВГ; Классен С., Олланд С., Бергер Дж. М. (сентябрь 2003 г.). «Структура области АТФазы топоизомеразы II и ее механизм ингибирования химиотерапевтическим агентом ICRF-187». Труды Национальной академии наук Соединенных Штатов Америки. 100 (19): 10629–34. Bibcode:2003PNAS..10010629C. Дои:10.1073 / pnas.1832879100. ЧВК 196855. PMID 12963818.
- ^ Вэй Х, Рутенбург А.Дж., Бечис СК, Вердин Г.Л. (ноябрь 2005 г.). «Нуклеотид-зависимое перемещение домена в АТФазном домене топоизомеразы ДНК типа IIA человека». Журнал биологической химии. 280 (44): 37041–7. Дои:10.1074 / jbc.M506520200. PMID 16100112. S2CID 35186716.
- ^ Аравинд Л., Лейпе Д.Д., Кунин Е.В. (сентябрь 1998 г.). «Топрим - консервативный каталитический домен в топоизомеразах типа IA и II, примазах типа DnaG, нуклеазах семейства OLD и белках RecR». Исследования нуклеиновых кислот. 26 (18): 4205–13. Дои:10.1093 / nar / 26.18.4205. ЧВК 147817. PMID 9722641.
- ^ PDB: 1BGW; Бергер Дж. М., Гамблин С. Дж., Харрисон С. К., Ван Дж. К. (январь 1996 г.). «Структура и механизм ДНК-топоизомеразы II». Природа. 379 (6562): 225–32. Bibcode:1996Натура 379..225Б. Дои:10.1038 / 379225a0. PMID 8538787. S2CID 4360011.
- ^ PDB: 1AB4; Мораис Кабрал Дж. Х., Джексон А. П., Смит К. В., Шикотра Н., Максвелл А., Лиддингтон Р. К. (август 1997 г.). «Кристаллическая структура домена разрыва-воссоединения ДНК-гиразы». Природа. 388 (6645): 903–6. Bibcode:1997Натура.388..903М. Дои:10.1038/42294. PMID 9278055. S2CID 4320715.
- ^ PDB: 1BJT; Фасс Д., Богден С.Э., Бергер Дж. М. (апрель 1999 г.). «Четвертичные изменения в топоизомеразе II могут направлять ортогональное движение двух цепей ДНК». Структурная биология природы. 6 (4): 322–6. Дои:10.1038/7556. PMID 10201398. S2CID 947461.
- ^ PDB: 2RGR; Донг К.С., Бергер Дж. М. (декабрь 2007 г.). «Структурная основа для распознавания ворот-ДНК и изгиба топоизомеразами типа IIA». Природа. 450 (7173): 1201–5. Bibcode:2007Натура.450.1201D. Дои:10.1038 / природа06396. PMID 18097402. S2CID 1756317.
- ^ Ванден Брок А., Лотц С., Ортис Дж., Ламур В. (октябрь 2019 г.). «Крио-ЭМ структура полного комплекса нуклеопротеидов ДНК-гиразы E. coli». Nature Communications. 10 (1): 4935. Bibcode:2019НатКо..10.4935V. Дои:10.1038 / s41467-019-12914-у. ЧВК 6821735. PMID 31666516.
- ^ PDB: 1ГУ; Корбетт К.Д., Шульцабергер Р.К., Бергер Дж.М. (май 2004 г.). «С-концевой домен ДНК-гиразы А принимает изгибающуюся ДНК складку бета-вертушки». Труды Национальной академии наук Соединенных Штатов Америки. 101 (19): 7293–8. Bibcode:2004PNAS..101.7293C. Дои:10.1073 / pnas.0401595101. ЧВК 409912. PMID 15123801.
- ^ PDB: 1MU5; Корбетт К.Д., Бергер Дж. М. (январь 2003 г.). «Структура субъединицы топоизомеразы VI-B: значение для механизма и эволюции топоизомеразы типа II». Журнал EMBO. 22 (1): 151–63. Дои:10.1093 / emboj / cdg008. ЧВК 140052. PMID 12505993.
- ^ Бергерат А., де Масси Б., Гадель Д., Варутас П.С., Николас А., Фортер П. (март 1997 г.). «Атипичная топоизомераза II из архей с последствиями для мейотической рекомбинации». Природа. 386 (6623): 414–7. Bibcode:1997Натура.386..414Б. Дои:10.1038 / 386414a0. PMID 9121560. S2CID 4327493.
- ^ PDB: 2Q2E; Корбетт К.Д., Бенедетти П., Бергер Дж. М. (июль 2007 г.). «Сборка голоэнзима и АТФ-опосредованная конформационная динамика топоизомеразы VI». Структурная и молекулярная биология природы. 14 (7): 611–9. Дои:10.1038 / nsmb1264. PMID 17603498. S2CID 2159631.
- ^ Рока Дж., Ван Дж. К. (май 1994 г.). «Транспортировка ДНК с помощью топоизомеразы ДНК типа II: свидетельство в пользу механизма с двумя воротами». Клетка. 77 (4): 609–16. Дои:10.1016/0092-8674(94)90222-4. PMID 8187179. S2CID 19776252.
- ^ Бергер Дж. М., Ван Дж. К. (февраль 1996 г.). «Последние разработки в структуре и механизме ДНК-топоизомеразы II». Текущее мнение в структурной биологии. 6 (1): 84–90. Дои:10.1016 / s0959-440x (96) 80099-6. PMID 8696977.
- ^ Альбертс Б. (18 ноября 2014 г.). Молекулярная биология клетки (Шестое изд.). Нью-Йорк, штат Нью-Йорк. ISBN 978-0-8153-4432-2. OCLC 887605755.
- ^ Уиллмор Э., де Ко С., Сантер Нью-Джерси, Тилби М.Дж., Джексон Г.Х., Остин, Калифорния, Дуркач, Б.В. (июнь 2004 г.). «Новый ингибитор ДНК-зависимой протеинкиназы, NU7026, усиливает цитотоксичность ядов топоизомеразы II, используемых при лечении лейкемии». Кровь. 103 (12): 4659–65. Дои:10.1182 / кровь-2003-07-2527. PMID 15010369.
- ^ Лю Л.Ф., Лю CC, Альбертс Б.М. (октябрь 1979 г.). «ДНК-топоизомераза Т4: новый АТФ-зависимый фермент, необходимый для инициации репликации ДНК бактериофага Т4». Природа. 281 (5731): 456–61. Bibcode:1979Натура.281..456л. Дои:10.1038 / 281456a0. PMID 226889. S2CID 4343962.
- ^ Стетлер Г.Л., Кинг Г.Дж., Хуанг В.М. (август 1979 г.). «Белки задержки ДНК Т4, необходимые для специфической репликации ДНК, образуют комплекс, обладающий АТФ-зависимой ДНК-топоизомеразной активностью». Труды Национальной академии наук Соединенных Штатов Америки. 76 (8): 3737–41. Bibcode:1979PNAS ... 76.3737S. Дои:10.1073 / pnas.76.8.3737. ЧВК 383908. PMID 226976.
- ^ а б Маккарти Д. (январь 1979 г.). «Гираза-зависимая инициация репликации ДНК бактериофага Т4: взаимодействия гиразы Escherichia coli с новобиоцином, кумермицином и продуктами гена задержки ДНК фага». Журнал молекулярной биологии. 127 (3): 265–83. Дои:10.1016/0022-2836(79)90329-2. PMID 372540.
- ^ Хуан WM (сентябрь 1986 г.). «52-белковая субъединица ДНК-топоизомеразы Т4 гомологична gyrA-белку гиразы». Исследования нуклеиновых кислот. 14 (18): 7379–90. ЧВК 311757. PMID 3020513.
- ^ Хуан WM (октябрь 1986 г.). «Нуклеотидная последовательность гена топоизомеразы ДНК типа II. Ген T4 бактериофага 39». Исследования нуклеиновых кислот. 14 (19): 7751–65. Дои:10.1093 / nar / 14.19.7751. ЧВК 311794. PMID 3022233.
- ^ Маккарти Д., Миннер С., Бернштейн Н., Бернштейн С. (октябрь 1976 г.). «Скорость удлинения ДНК и распределение точек роста фага Т4 дикого типа и янтарного мутанта с задержкой ДНК». Журнал молекулярной биологии. 106 (4): 963–81. Дои:10.1016/0022-2836(76)90346-6. PMID 789903.
- ^ Муфтий С., Бернштейн Х (октябрь 1974 г.). «Задерживающие ДНК мутанты бактериофага Т4». Журнал вирусологии. 14 (4): 860–71. Дои:10.1128 / JVI.14.4.860-871.1974. ЧВК 355592. PMID 4609406.
- ^ Хайман П. (август 1993 г.). «Генетика эффекта Луриа-Латарджета в бактериофаге T4: доказательства участия множественных путей репарации ДНК». Генетические исследования. 62 (1): 1–9. Дои:10,1017 / с0016672300031499. PMID 8405988.
- ^ Miskimins R, Schneider S, Johns V, Bernstein H (июнь 1982). «Участие топоизомеразы в реактивации множественности фага Т4». Генетика. 101 (2): 157–77. ЧВК 1201854. PMID 6293912.
- ^ Кассуто Э (сентябрь 1984 г.). «Образование ковалентно замкнутой гетеродуплексной ДНК за счет комбинированного действия гиразы и белка RecA». Журнал EMBO. 3 (9): 2159–64. Дои:10.1002 / j.1460-2075.1984.tb02106.x. ЧВК 557658. PMID 6092061.
дальнейшее чтение
внешняя ссылка
- ДНК + топоизомеразы, + тип + II в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)