EGF-подобный домен - EGF-like domain
Эта статья отсутствует информация о консенсусном цистиновом повторе, положении дифульфидной связи (остатки, внутри / внутри?). (Март 2019 г.) |
| EGF-подобный домен | |||||||||
|---|---|---|---|---|---|---|---|---|---|
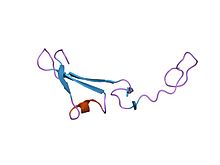 Структура домена херегулина-альфа, подобного эпидермальному фактору роста.[1] | |||||||||
| Идентификаторы | |||||||||
| Символ | EGF | ||||||||
| Pfam | PF00008 | ||||||||
| Pfam клан | CL0001 | ||||||||
| ИнтерПро | IPR000742 | ||||||||
| PROSITE | PDOC00021 | ||||||||
| SCOP2 | 1апо / Объем / СУПФАМ | ||||||||
| CDD | cd00053 | ||||||||
| |||||||||
| EGF-подобный домен, внеклеточный | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 кристаллическая структура внеклеточного сегмента интегрина alphavbeta3 | |||||||||
| Идентификаторы | |||||||||
| Символ | EGF_2 | ||||||||
| Pfam | PF07974 | ||||||||
| Pfam клан | CL0001 | ||||||||
| ИнтерПро | IPR013111 | ||||||||
| CDD | cd00054 | ||||||||
| |||||||||
В EGF-подобный домен это эволюционно сохраненный белковый домен, который получил свое название от фактор роста эпидермиса где это было впервые описано. Он состоит от 30 до 40 аминокислотных остатков и обнаружен в большом количестве белков, в основном животных.[2][3] Большинство случаев EGF-подобного домена обнаруживается во внеклеточном домене мембраносвязанные белки или в белках, которые известны как секретный. Исключением является простагландин-эндопероксидсинтаза. EGF-подобный домен включает 6 цистеин остатки, которые в эпидермальном факторе роста образуют 3 дисульфидные связи. Структура 4-дисульфидных EGF-доменов решена из ламинин и интегрин белки. Основная структура EGF-подобных доменов представляет собой двухцепочечный β-лист с последующей петлей к короткому С-концу двухцепочечного β-листа. Эти два β-листа обычно обозначают как основной (N-концевой) и вспомогательный (C-концевой) листы.[4] EGF-подобные домены часто встречаются в многочисленных тандемных копиях в белках: обычно эти повторы складывать вместе, чтобы сформировать единый линейный соленоидный домен блок как функциональная единица.
Подтипы
Несмотря на сходство EGF-подобных доменов, были идентифицированы различные подтипы доменов.[5] Двумя основными предлагаемыми типами EGF-подобных доменов являются EGF-подобный домен человека (hEGF) и C1r-подобный домен комплемента (cEGF),[4] который был впервые идентифицирован в протеазе комплемента человека C1r.[5] C1r - очень специфичный сериновая протеаза инициируя классический путь активация комплемента во время иммунного ответа.[6] Как hEGF-, так и cEGF-подобные домены содержат три дисульфида и происходят от общего предка, несущего четыре дисульфида, один из которых был утерян в процессе эволюции. Кроме того, cEGF-подобные домены можно разделить на два подтипа (1 и 2), тогда как все hEGF-подобные домены принадлежат к одному подтипу.[4]
Дифференциация cEGF-подобных и hEGF-подобных доменов и их подтипов основана на структурных особенностях и связанности их дисульфидных связей. cEGF- и hEGF-подобные домены имеют различную форму и ориентацию минорного листа, а один C-концевой полуцистин имеет другое положение. Утерянные цистеины общего предка различаются между cEGF- и hEGF-подобными доменами, и, следовательно, эти типы различаются по своим дисульфидным связям. Дифференциация cEGF на подтипы 1 и 2, которая, вероятно, произошла после его расщепления от hEGF, основана на различном количестве остатков между отдельными полуцистинами. Расположенный на N-конце кальций-связывающий мотив можно найти как в hEGF-, так и в cEGF-подобных доменах, и поэтому он не подходит для их разделения.[4]
hEGF- и cEGF-подобные домены также содержат посттрансляционные модификации, которые часто необычны и различаются между hEGF- и cEGF-подобными доменами. Эти посттрансляционные модификации включают O-гликозилирование, в основном модификации O-фукозы и β-гидроксилирование аспартата и остатков аспарагина. Модификации O-фукозы были обнаружены только в hEGF-подобных доменах, и они важны для правильной укладки hEGF-подобного домена. β-Гидроксилирование проявляется в hEGF- и cEGF-подобных доменах, первый гидроксилируется по аспарагиновой кислоте, а второй гидроксилируется по остатку аспарагина. Биологическая роль этой посттрансляционной модификации неясна.[4] но у мышей с нокаутом фермента аспартил-β-гидроксилирования обнаруживаются дефекты развития.[7]
Белки, содержащие EGF-подобные домены, широко распространены и могут быть исключительно hEGF- или cEGF-подобными или содержать их смесь. Во многих митогенных белках и белках развития, таких как Notch и Delta, EGF-подобные домены относятся только к типу hEGF. Другие белки содержат только cEGF, такие как тромбомодулин и ЛПНП-рецептор. В смешанных EGF-белках hEGF- и cEGF-подобные домены сгруппированы вместе, причем hEGF всегда являются N-концевыми по отношению к cEGF. Такие белки участвуют в свертывании крови или являются компонентами внеклеточного матрикса, например фибриллин и LTBP-1 (белок 1, связывающий бета-фактор роста с латентным трансформирующим фактором роста). В дополнение к вышеупомянутым трем дисульфидным hEGF- и cEGF-подобным типам существуют белки, несущие четырехдисульфидный EGF-подобный домен, такой как ламинин и интегрин.[4]
Два основных подтипа EGF-подобных доменов hEGF и cEGF не только различаются по своей структуре и конформации, но также имеют разные функции. Эта гипотеза подтверждается исследованиями на LTBP-1. LTBP-1 закрепляет трансформирующий фактор роста β (TGF-β) во внеклеточном матриксе. hEGF-подобные домены играют роль в нацеливании сборки LTBP-1 / TGF-β на внеклеточный матрикс. Прикрепившись к внеклеточному матриксу, TGF-β диссоциирует от субъединиц hEGF, чтобы обеспечить его последующую активацию. cEGF-подобные домены, по-видимому, играют неспецифическую роль в этой активации, способствуя отщеплению LTBP-1 от TGF-β различными протеазами.[4]
В заключение, хотя отдельные EGF-подобные домены сгруппированы, подтипы можно четко разделить по их последовательности, конформации и, что наиболее важно, их функции.
Роль в иммунной системе и апоптозе
Selectins, группа белков, которые участвуют в лейкоциты катящиеся к источнику воспаления, содержат EGF-подобный домен вместе с лектиновым доменом и короткие консенсусные повторы (SCR).[8][9] Функции EGF-подобного домена различаются для разных типов селектина. Канзас и соавторы смогли показать, что EGF-подобный домен не требуется для максимальной клеточной адгезии в L-селектине (выраженном на лимфоцитах). Однако он участвует как в распознавании лиганда, так и в адгезии Р-селектина (экспрессируется на тромбоцитах), а также может участвовать во взаимодействиях белок-белок. Было высказано предположение, что взаимодействия между лектиновыми доменами и углеводными лигандами могут быть кальций-зависимыми.[8]
Незрелый человек дендритные клетки по-видимому, требуется взаимодействие с EGF-подобными доменами селектинов во время процесса их созревания. Блокирование этого взаимодействия с моноклональными антителами к EGF-подобному домену предотвращает созревание дендритных клеток. Незрелые клетки не могут активировать Т-клетки и вырабатывать меньше интерлейкина 12, чем дендритные клетки дикого типа.[10]
Фан и другие. может показать, что искусственное введение Сайт N-гликозилирования в EGF-подобные домены в P- и L-селектинах увеличивали сродство селектинов к их лигандам и приводили к более медленному скатыванию.[9] Следовательно, EGF-подобные домены, по-видимому, играют решающую роль в перемещении лейкоцитов в направлении воспалительных стимулов.
EGF-подобный домен также является частью ламининов, важной группы внеклеточных белков. EGF-подобные домены обычно маскируются в неповрежденных мембранах, но становятся доступными, когда мембрана разрушается, например во время воспаления, тем самым стимулируя рост мембраны и восстанавливая поврежденные части мембраны.[11]
Более того, было показано, что повторы EGF-подобного домена домена стабилизина-2 специфически распознают и связывают апоптотические клетки, вероятно, путем распознавания фосфатидилсерин, маркер апоптотических клеток («съешь меня-сигнал»).[12] Парк и другие. далее продемонстрировали, что домены способны конкурентно ухудшать распознавание апоптотических клеток макрофагами.
В заключение, EGF-подобный домен, по-видимому, играет жизненно важную роль в иммунных ответах, а также в устранении мертвых клеток в организме.
Связывающий кальций
Кальций-связывающие EGF-подобные домены (cbEGF-подобные домены) играют важную роль в таких заболеваниях, как Синдром Марфана[13] или геморрагическое заболевание, связанное с Х-хромосомой гемофилия B [14] и являются одними из самых распространенных внеклеточных кальций-связывающих доменов.[15] Важно отметить, что cbEGF-подобные домены придают определенные функции множеству белков в каскаде свертывания крови. Примеры включают факторы свертывания крови. VII, IX и Икс, протеин C и его кофактор протеин S.[15]
Кальций-связывающие EGF-подобные домены обычно состоят из 45 аминокислот, расположенных в виде двух антипараллельных бета-листов.[15] Несколько остатков цистеина в этой последовательности образуют дисульфидные мостики.
cbEGF-подобные домены не обнаруживают значительных структурных отклонений от EGF-подобных доменов; однако, как следует из названия, cbEGF-подобные домены связывают один ион кальция. Аффинность связывания с кальцием широко варьирует и часто зависит от соседних доменов.[15] Консенсусным мотивом связывания кальция является Asp-Leu / Ile-Asp-Gln-Cys. Координация кальция сильно коррелирует с необычной посттрансляционной модификацией cbEGF-подобных доменов: либо аспарагин, либо аспартат бета-гидроксилированы, давая начало эритро-бета-гидроксиаспарагину (Hyn) или эритро-бета-гидроксиаспарагиновой кислоте (Hya) соответственно. Hya можно найти в N-концевом модуле cbEGF (см. Ниже) факторов IX, X и белка C. Модификация Hyn, по-видимому, более распространена, чем Hya, и, как было показано, встречается в фибриллине-1, белке внеклеточного матрикса. .[16] Обе модификации катализируются диоксигеназой Asp / Asn-бета-гидроксилазой,[17] и уникальны для доменов EGF у эукариот.[15]
Сообщалось о дальнейших посттрансляционных модификациях. Гликозилирование в форме О-связанных ди- или трисахаридов может происходить по остатку серина между первыми двумя цистеинами факторов свертывания крови VII и IX.[18][19][20] Фактор VII демонстрирует O-связанную фукозу по Ser60.[20]
Множественные домены cbEGF часто соединяются одной или двумя аминокислотами с образованием более крупных повторяющихся массивов, называемых здесь «модулями cbEGF». В каскаде свертывания крови факторы свертывания крови VII, IX и X и протеин C содержат тандем из двух модулей cbEGF, тогда как протеин S - четыре. Поразительно, но в фибриллине-1 и фибриллине-2 было обнаружено 43 модуля cbEGF.[21] Модульность этих белков усложняет взаимодействие белок-белок, но также и модульное взаимодействие. В факторах VII, IX и X двум модулям cbEGF предшествует N-концевой модуль, содержащий гамма-карбоксиглутаминовую кислоту (Gla) ( Модуль Gla ).[15] Исследования in vitro тандема Gla-cbEGF, выделенного из фактора X, выявили Kd-значение 0,1 мМ для связывания кальция [18] при концентрации свободного кальция в плазме крови примерно 1,2 мМ. Удивительно, но в отсутствие модуля Gla модуль cbEGF демонстрирует Kd-значение 2,2 мМ для кальция.[17] Таким образом, наличие модуля Gla увеличивает сродство к кальцию в 20 раз. Точно так же активность модулей Gla и сериновой протеазы модифицируется модулями cbEGF. В отсутствие кальция модули Gla и cbEGF очень подвижны. Однако, поскольку модуль cbEGF ассоциируется с кальцием, перемещение модуля Gla значительно ограничено, поскольку модуль cbEGF теперь принимает конформацию, которая фиксирует соседний модуль Gla в фиксированном положении.[22][23] Следовательно, координация кальция вызывает конформационные изменения, которые, в свою очередь, могут модулировать ферментативную активность.
Нарушение координации кальция может привести к серьезным нарушениям. Нарушение связывания кальция с фактором свертывания IX способствует развитию гемофилии B. У лиц, страдающих этим наследственным заболеванием, обычно развиваются кровоизлияния, потенциально приводящие к опасным для жизни состояниям. Причина гемофилии B - снижение активности или дефицит фактора свертывания крови IX. Считается, что точечные мутации, приводящие к снижению сродства фактора IX к кальцию, связаны с этим нарушением свертываемости крови.[15] На молекулярной основе кажется, что гемофилия B может быть результатом нарушения способности эффективно локализовать модуль Gla, как это обычно происходит после координации кальция модулем cbEGF в полностью функциональном факторе IX.[15] Считается, что этот дефект нарушает биологическую функцию фактора IX. Аналогичная проблема возникает у пациентов, страдающих гемофилией B и несущих мутацию (Glu78Lys) в факторе IX, которая предотвращает взаимодействие двух модулей cbEGF друг с другом.[15] Напротив, у здоровых людей Glu78 в первом модуле cbEGF связывается с Arg94 во втором модуле cbEGF и тем самым выравнивает оба модуля.[24] Таким образом, взаимодействия домен-домен (частично облегченные координацией кальция) имеют решающее значение для каталитической активности белков, участвующих в каскаде свертывания крови.
Белки, содержащие этот домен
Ниже приведен список белков человека, содержащих EGF-подобный домен:
- AGC1; УСМЕШКА; AREG; ATRN; ATRNL1;
- BCAN; BMP1; BTC;
- C1S; CASPR4; CD248; CD93; CELSR1; CELSR2; CELSR3; CLEC14A; CNTNAP1; CNTNAP2; CNTNAP3; CNTNAP4; CNTNAP5; КОМП; СОХ-2; CRB1; CRB2; CSPG3; CUBN;
- DLK1; DLL1; DLL3; DLL4; DNER;
- EDIL3; EGF; EGFL11; EGFL8; EGFL9; EGFLAM; EPGN; EREG;
- F7; F9; F10; F12; ТОЛСТЫЙ; FAT2; FAT4; FBN1; FBN2; FBN3;
- ГАЗ6;
- HABP2; HBEGF; HEG1; HGFAC; HMCN1; HSPG2;
- ITGB5;
- JAG1; JAG2;
- LDLR; LRP1; LRP10; LRP1B; LRP2; LRP4; LRP5; LRP6; LRP8; LTBP1; LTBP2; LTBP3; LTBP4;
- MATN1; MATN2; MATN3; MATN4; MEGF12; MEGF6; MEP1A; MEP1B; MFGE8; MMRN1; MMRN1; MUC4;
- НАГПА; NID1; NID2; NOTCH1; NOTCH2; NOTCH2NL; NOTCH3; NOTCH4; NRG1; NRG2; NRG3; NRG4; NRXN1; NRXN2; NRXN3; NTNG2;
- ODZ1; ODZ2; OIT3;
- ПЛАТ; PP187; PROC; PROS1; ПРОЗ; PTGS1; PTGS2;
- RAMP;
- SCUBE1; SCUBE2; SCUBE3; SEL-OB; SELE; ПРОДАВАТЬ; SELP; SLIT1; SLIT2; SLIT3; SNED1; STAB1; STAB2; SVEP1;
- TECTA; TGFA; THBD; THBS1; THBS2; THBS4; TIE1; TLL1; TLL2; TMEFF1; TMEFF2; TNC; TNXB;
- UMOD;
- ВАСН; VCAN; VLDLR; VWA2;
- WIF1;
- ZAN;
Смотрите также
Рекомендации
- ^ Нагата К., Кохда Д., Хатанака Х. и др. (Август 1994 г.). «Структура раствора подобного эпидермальному фактору роста домена херегулина-альфа, лиганда p180erbB-4». EMBO J. 13 (15): 3517–23. Дои:10.1002 / j.1460-2075.1994.tb06658.x. ЧВК 395255. PMID 8062828.
- ^ Даунинг А.К., Нотт В., Вернер Дж. М., Карди С. М., Кэмпбелл И. Д., Хэндфорд, Пенсильвания (май 1996 г.). «Структура решения пары кальций-связывающих эпидермальных факторов роста подобных доменов: последствия для синдрома Марфана и других генетических нарушений». Клетка. 85 (4): 597–605. Дои:10.1016 / S0092-8674 (00) 81259-3. PMID 8653794. S2CID 15410014.
- ^ Борк П., Даунинг А. К., Киффер Б., Кэмпбелл И. Д. (май 1996 г.). «Структура и распределение модулей во внеклеточных белках». Q. Rev. Biophys. 29 (2): 119–67. Дои:10.1017 / S0033583500005783. PMID 8870072.
- ^ а б c d е ж грамм Воутерс MA, Rigoutsos I, Chu CK, Feng LL, Sparrow DB, Dunwoodie SL (2005). «Эволюция отдельных доменов EGF со специфическими функциями». Белковая наука. 14 (4): 1091–103. Дои:10.1110 / пс. 041207005. ЧВК 2253431. PMID 15772310.
- ^ а б Берш Б., Эрнандес Дж. Ф., Марион Д., Арлауд Дж. Дж. (1998). «Структура раствора эпидермального фактора роста (EGF) -подобного модуля протеазы комплемента C1r человека, атипичного члена семейства EGF». Биохимия. 37 (5): 1204–14. Дои:10.1021 / bi971851v. PMID 9477945.
- ^ Circolo A, Гарнье Дж., Воланакис Дж. Э. (2003). «Новый мышиный ген, связанный с комплементом, кодирующий C1r-подобный сывороточный белок». Молекулярная иммунология. 39 (14): 899–906. Дои:10.1016 / S0161-5890 (02) 00283-3. PMID 12686506.
- ^ Стенфло Дж., Олин А.К., Оуэн В.Г., Шнайдер В.Дж. (1988). «Бета-гидроксиаспарагиновая кислота или бета-гидроксиаспарагин в рецепторах липопротеинов низкой плотности крупного рогатого скота и в тромбомодулине крупного рогатого скота». Журнал биологической химии. 263 (1): 21–24. PMID 2826439.
- ^ а б Канзас Г.С., Сондерс КБ, Лей К. и др. (1994). «Роль домена P-селектина, подобного эпидермальному фактору роста, в распознавании лиганда и клеточной адгезии». J Cell Biol. 124 (4): 609–18. Дои:10.1083 / jcb.124.4.609. ЧВК 2119911. PMID 7508943.
- ^ а б Phan UT, Waldron TT, Springer TA (2006). «Ремоделирование интерфейса лектин-EGF-подобный домен в P- и L-селектине увеличивает адгезионную способность и сопротивление сдвигу под действием гидродинамической силы». Нат Иммунол. 7 (8): 883–9. Дои:10.1038 / ni1366. ЧВК 1764822. PMID 16845394.
- ^ Чжоу Т., Чжан Ю., Сунь Г. и др. (2006). «Моноклональное антитело против лектина P-селектина и домена EGF ингибирует созревание незрелых дендритных клеток человека». Опыт Мол Патол. 80 (2): 171–6. Дои:10.1016 / j.yexmp.2005.10.004. PMID 16413535.
- ^ Löffler, G; Петридес, ЧП; Генрих, ПК (1997). Биохимия и патобиохимия (5-е изд.). Берлин, Гейдельберг: Springer-Verlag. п. 747. ISBN 3-540-59006-4.
- ^ Пак С.И., Ким С.И., Юнг М.Ю. и др. (2008). «Повторение домена, подобного эпидермальному фактору роста, табилина-2 распознает фосфатидилсерин во время очищения от трупа клеток». Mol Cell Biol. 28 (17): 5288–98. Дои:10.1128 / MCB.01993-07. ЧВК 2519725. PMID 18573870.
- ^ Хэндфорд PA, Даунинг AK, Rao Z, Hewett DR, Sykes BC, Kielty CM (1991). «Свойства связывания кальция и молекулярная организация доменов, подобных эпидермальному фактору роста, в человеческом фибриллине-1». J. Biol. Chem. 270 (12): 6751–6. Дои:10.1074 / jbc.270.12.6751. PMID 7896820.
- ^ Handford PA, Mayhew M, Baron M, Winship PR, Campbell ID, Brownlee GG (1991). «Ключевые остатки, участвующие в кальций-связывающих мотивах в EGF-подобных доменах». Природа. 351 (6322): 164–7. Дои:10.1038 / 351164a0. PMID 2030732. S2CID 4338236.
- ^ а б c d е ж грамм час я Стенфло Дж, Стенберг Й, Мураньи А (2000). «Кальций-связывающие EGF-подобные модули в протеиназах свертывания: функция иона кальция во взаимодействиях модулей». Biochimica et Biophysica Acta (BBA) - Структура белка и молекулярная энзимология. 1477 (1–2): 51–63. Дои:10.1016 / s0167-4838 (99) 00262-9. PMID 10708848.
- ^ Гланвилл Р.В., Циан Р.К., МакКлюр Д.В., Маслен К.Л. и др. (1994). «Связывание кальция, гидроксилирование и гликозилирование предшественников эпидермального фактора роста-подобных доменов фибриллина-1, белка гена Марфана». J. Biol. Chem. 269 (43): 26630–4. PMID 7929395.
- ^ а б Jia S, VanDusen WJ, Diehl RE и др. (1992). «Клонирование кДНК и экспрессия бычьей аспартил (аспарагинил) бета-гидроксилазы». J. Biol. Chem. 267 (20): 14322–7. PMID 1378441.
- ^ а б Valcarce C, Selander-Sunnerhagen M, Tämlitz AM, Drakenberg T, Björk I, Stenflo J (1996). «Сродство к кальцию NH2-терминального эпидермального фактора роста, подобного модулю фактора X». J. Biol. Chem. 268 (35): 26673–8. PMID 8253800.
- ^ Нисимура Х., Кавабата С., Кисиэль В. и др. (1989). «Идентификация дисахарида (Xyl-Glc) и трисахарида (Xyl2-Glc) O-гликозидно связанных с остатком серина в первом эпидермальном факторе роста-подобном домене человеческих факторов VII и IX и протеина Z и бычьего протеина Z». J. Biol. Chem. 264 (34): 20320–5. PMID 2511201.
- ^ а б Бьорн С., Фостер Д., Тим Л. и др. (1991). «Плазма человека и рекомбинантный фактор VII». J. Biol. Chem. 266 (17): 11051–7. PMID 1904059.
- ^ Пиха-Госсак А., Соссин В., Рейнхардт Д. Т. и др. (2012). «Эволюция внеклеточных фибриллинов и их функциональных доменов». PLOS ONE. 7 (3): 33560. Дои:10.1371 / journal.pone.0033560. ЧВК 3306419. PMID 22438950.
- ^ Саннерхаген М., Форсен С., Хоффрен А., Дракенберг Т., Телеман О., Стенфло Дж. (1995). «Структура домена Gla, свободного от Ca (2 +), проливает свет на связывание белков свертывания крови с мембраной». Структурная и молекулярная биология природы. 2 (6): 504–9. Дои:10.1038 / nsb0695-504. PMID 7664114. S2CID 8570806.
- ^ Sunnerhagen M, Olah GA, Stenflo J, Forsen S, Drakenberg T, Trewhella J (1996). «Относительная ориентация доменов Gla и EGF в факторе X коагуляции изменяется из-за связывания Ca2 + с первым доменом EGF. Комбинированное исследование ЯМР-малоуглового рентгеновского рассеяния». Биохимия. 35 (36): 11547–59. Дои:10.1021 / bi960633j. PMID 8794734.
- ^ Кристоф О.Д., Лентинг П.Дж., Колкман Дж.А., Браунли Г.Г., Мертенс К. (1988). «Остатки фактора свертывания крови IX Glu78 и Arg94 обеспечивают связь между обоими доменами, подобными эпидермальному фактору роста, которые имеют решающее значение во взаимодействии с легкой цепью фактора VIII». J. Biol. Chem. 273 (1): 222–27. Дои:10.1074 / jbc.273.1.222. PMID 9417068.