Рецептор AMPA - AMPA receptor

В Рецептор α-амино-3-гидрокси-5-метил-4-изоксазолепропионовой кислоты (также известен как AMPA рецептор, AMPAR, или квисквалатный рецептор) является ионотропный трансмембранный рецептор для глутамат это быстро посредничает синаптический передача в Центральная нервная система (ЦНС). Он традиционно классифицируется как неNMDA -типа рецептора, наряду с каинатный рецептор. Его название происходит от его способности активироваться искусственным аналогом глутамата. AMPA. Рецептор был впервые назван «рецептором quisqualate» Уоткинсом и его коллегами в честь природного агониста. quisqualate и только позже получил название «рецептор AMPA» в честь селективного агониста, разработанного Таге Оноре и его коллегами из Королевской датской фармацевтической школы в Копенгагене.[1] AMPAR можно найти во многих частях мозг и являются наиболее часто встречающимся рецептором в нервная система. Кодируемое GRIA2 ядро связывания лиганда рецептора AMPA (см. Ниже) было первым ионным каналом рецептора глутамата, который кристаллизованный.[2]
Структура и функции
Субъединичный состав
AMPAR состоят из четырех типов субъединиц, кодируемых разными генами, обозначенных как GRIA1 (также называемый GluA1 или GluR1), GRIA2 (также называемый GluA2 или GluR2), GRIA3 (Также называется GluA3 или GluR3), и GRIA4 (также называемые GluA4 или GluRA-D2), которые в совокупности образуют тетрамеры.[3][4][5] Большинство AMPAR являются гетеротетрамерными, состоящими из симметричного «димера димеров» GluA2 или GluA1, GluA3 или GluA4.[6][7] Димеризация начинается в эндоплазматический ретикулум при взаимодействии N-концевых доменов LIVBP, затем «застревает» через лиганд-связывающий домен в трансмембранную ионную пору.[7]
Конформация субъединичного белка в плазматическая мембрана вызывало споры некоторое время. В то время как аминокислотная последовательность субъединицы указывает на то, что, по-видимому, существует четыре трансмембранных домена (части белка, которые проходят через плазматическую мембрану), белки, взаимодействующие с субъединицей, указывают на то, что N-конец казались внеклеточными, а C-конец казалось внутриклеточным. Однако, если бы каждый из четырех трансмембранных доменов пошел весь путь через плазматическую мембрану, тогда два конца должны быть на одной стороне мембраны. В конце концов было обнаружено, что второй «трансмембранный» домен на самом деле вообще не пересекает мембрану, а изгибается внутри мембраны и возвращается во внутриклеточную сторону.[8] Когда четыре субъединицы тетрамера объединяются, этот второй мембранный домен образует проницаемую для ионов пору рецептора.
Субъединицы AMPAR больше всего различаются по своей С-концевой последовательности, которая определяет их взаимодействие с каркасными белками. Все AMPAR содержат PDZ-связывающие домены, но которые PDZ домен они связаны с разными. Например, GluA1 связывается с SAP97 через домен PDZ класса I SAP97,[9] в то время как GluA2 связывается с PICK1[10] и GRIP / ABP. Следует отметить, что AMPAR не могут напрямую связываться с общим синаптическим белком. PSD-95 из-за несовместимости доменов PDZ, хотя они взаимодействуют с PSD-95 через звездочет (прототипный член семейства TARP вспомогательных субъединиц AMPAR).[11]
Фосфорилирование AMPAR могут регулировать локализацию канала, проводимость и вероятность открытия. GluA1 имеет четыре известных сайта фосфорилирования в серин 818 (S818), S831, треонин 840 и S845 (другие субъединицы имеют аналогичные сайты фосфорилирования, но GluR1 изучен наиболее полно). S818 фосфорилируется протеинкиназа C, и необходимо для долгосрочное потенцирование (LTP; роль GluA1 в LTP см. Ниже).[12] S831 фосфорилируется CaMKII и PKC во время LTP, что помогает доставить GluA1-содержащий AMPAR к синапс,[13] и увеличивает их одноканальную проводимость.[14] Сайт T840 был обнаружен совсем недавно и был вовлечен в LTD.[15] Наконец, S845 фосфорилируется PKA, которая регулирует вероятность его открытия.[16]
Функция ионного канала
У каждого AMPAR есть четыре сайта, на которые агонист (например, глутамат) могут связываться, по одному на каждую субъединицу.[6] Предполагается, что сайт связывания образован N-концевым хвостом и внеклеточной петлей между третьим и четвертым трансмембранными доменами.[17] Когда агонист связывается, эти две петли движутся навстречу друг другу, открывая поры. Канал открывается, когда заняты два сайта,[18] и увеличивает свой ток по мере того, как занято больше сайтов связывания.[19] После открытия канал может подвергнуться быстрой десенсибилизации, останавливая ток. Считается, что механизм десенсибилизации связан с небольшим изменением угла одной из частей сайта связывания, закрывающей поры.[20] AMPAR открываются и закрываются быстро (1 мс) и, таким образом, отвечают за большую часть быстрых возбуждающих сигналов. синаптическая передача в центральной нервной системе.[18] Проницаемость AMPAR для кальций и другие катионы, такие как натрий и калий, управляется субъединицей GluA2. Если в AMPAR отсутствует субъединица GluA2, то он будет проницаемым для натрия, калия и кальция. Присутствие субъединицы GluA2 почти всегда делает канал непроницаемым для кальция. Это определяется пост-транскрипционный модификация - Редактирование РНК - из Q -к-р сайт редактирования GluA2 мРНК. Вот, A → I редактирование изменяет незаряженный аминокислота глутамин (Q) к положительно заряженному аргинин (R) в ионном канале рецептора. Положительно заряженная аминокислота в критической точке делает энергетически невыгодным проникновение кальция в клетку через поры. Почти все субъединицы GluA2 в ЦНС преобразованы в форму GluA2 (R). Это означает, что основными ионами, управляемыми AMPAR, являются натрий и калий, что отличает AMPAR от Рецепторы NMDA (другие основные ионотропные рецепторы глутамата в головном мозге), которые также допускают приток кальция. Однако рецепторы AMPA и NMDA имеют равновесный потенциал около 0 мВ. Предполагается, что предотвращение проникновения кальция в клетку при активации GluA2-содержащих AMPARs предотвращает эксайтотоксичность.[21]
Субъединичный состав AMPAR также важен для модуляции этого рецептора. Если в AMPAR отсутствуют субъединицы GluA2, то он подвержен блокированию в зависимости от напряжения классом молекул, называемым полиамины. Таким образом, когда нейрон находится в деполяризованный с мембранным потенциалом полиамины будут сильнее блокировать канал AMPAR, предотвращая поток ионов калия через поры канала. Таким образом, считается, что AMPAR, лишенные GluA2, обладают внутренне исправляющим действием. Кривая I / V, что означает, что они пропускают наружный ток меньше, чем внутренний ток на эквивалентном расстоянии от реверсивного потенциала. Проницаемые для кальция AMPAR обычно обнаруживаются на ранних этапах постнатального развития, на некоторых интернейронах или в дофаминовых нейронах вентральная тегментальная область после воздействия наркотического средства, вызывающего привыкание.[22]
Наряду с Редактирование РНК, альтернативное сращивание позволяет использовать ряд функциональных субъединиц рецептора AMPA за пределами того, что кодируется в геном. Другими словами, хотя один ген (GRIA1–GRIA4) кодируется для каждой субъединицы (GluA1 – GluA4), сплайсинг после транскрипции из ДНК позволяет некоторым экзоны переводиться взаимозаменяемо, что приводит к появлению нескольких функционально различных субъединиц от каждого гена.
Последовательность флип / флоп является одним из таких взаимозаменяемых экзонов. Последовательность из 38 аминокислот, обнаруженная до (т. Е. До N-конец из) четвертый мембранный домен во всех четырех субъединицах AMPAR, он определяет скорость десенсибилизации[23] рецептора, а также скорость, с которой рецептор повторно сенсибилизируется[24] и скорость закрытия канала.[25] Перевернутая форма присутствует в пренатальных рецепторах AMPA и дает устойчивый ток в ответ на активацию глутамата.[26]
Синаптическая пластичность
Рецепторы AMPA (AMPAR) являются рецепторы глутамата и катион каналы которые являются неотъемлемой частью пластичность и синаптическая передача у многих постсинаптических мембран. Одна из наиболее широко и тщательно изученных форм пластичности нервной системы известна как долгосрочное потенцирование, или LTP. Есть два необходимых компонента LTP: пресинаптический глутамат релиз и постсинаптическая деполяризация. Следовательно, ДП может быть индуцирована экспериментально в парной электрофизиологическая запись когда пресинаптическая клетка стимулируется высвобождением глутамата на постсинаптической клетке, которая деполяризована. Типичный протокол индукции LTP включает «столбнячную» стимуляцию, то есть стимуляцию 100 Гц в течение 1 секунды. Когда кто-то применяет этот протокол к паре ячеек, вы увидите устойчивое увеличение амплитуды возбуждающий постсинаптический потенциал (ВПСП) после столбняка. Этот ответ интересен, поскольку считается, что он является физиологическим коррелятом обучения и памяти в клетке. Фактически, было показано, что, следуя парадигме единственного парного избегания у мышей, LTP может быть зарегистрирован у некоторых гиппокамп синапсы in vivo.[27]
Молекулярная основа LTP была тщательно изучена, и было показано, что AMPAR играют важную роль в этом процессе. И GluR1, и GluR2 играют важную роль в синаптической пластичности. Теперь известно, что физиологический коррелят, лежащий в основе увеличения размера ВПСП, - это постсинаптическая активация AMPAR на мембране,[28] что достигается за счет взаимодействия AMPAR со многими клеточными белками.
Самое простое объяснение LTP следующее (см. долгосрочное потенцирование статью для более подробного описания). Глутамат связывается с постсинаптическими AMPAR и другим рецептором глутамата, Рецептор NMDA (NMDAR). Связывание лиганда вызывает открытие AMPAR, и Na+ попадает в постсинаптическую клетку, вызывая деполяризацию. NMDAR, с другой стороны, не открываются напрямую, потому что их поры закрываются при мембранном потенциале покоя Mg.2+ ионы. NMDAR могут открываться только тогда, когда деполяризация от активации AMPAR приводит к отталкиванию Mg2+ катион во внеклеточное пространство, позволяя поре пропускать ток. Однако, в отличие от AMPAR, NMDAR проницаемы как для Na+ и Ca2+. CA2+ который проникает в клетку, запускает активацию AMPARs в мембране, что приводит к длительному увеличению размера EPSP, лежащего в основе LTP. Поступление кальция также фосфорилирует CaMKII, который фосфорилирует AMPAR, увеличивая их одноканальную проводимость.
Торговля рецепторами AMPA

Молекулярный и сигнальный ответ на стимулы, индуцирующие LTP
Механизм LTP уже давно является предметом дискуссий, но в последнее время механизмы пришли к определенному консенсусу. AMPAR играют ключевую роль в этом процессе, поскольку одним из ключевых индикаторов индукции LTP является увеличение отношения AMPAR к NMDAR после высокочастотной стимуляции. Идея состоит в том, что AMPAR передаются от дендрита в синапс и включаются через серию сигнальных каскадов.
AMPAR изначально регулируются на уровне транскрипции в своих 5’-промоторных областях. Существуют важные доказательства, указывающие на транскрипционный контроль рецепторов AMPA в долговременной памяти посредством белка, связывающего элемент ответа цАМФ (CREB ) и Митоген-активированные протеинкиназы (МАПК).[29] Сообщения переведены на черновой эндоплазматический ретикулум (черновая ER) и модифицированная там. Состав субъединиц определяется во время модификации на грубом ER.[10] После пост-ER обработки в аппарате Гольджи AMPAR высвобождаются в перисинаптическую мембрану в качестве резерва, ожидающего запуска процесса LTP.
Первым ключевым шагом в процессе связывания глутамата с NMDAR является приток кальция через рецепторы NMDA и, как следствие, активация Ca2+/ кальмодулин-зависимая протеинкиназа (CaMKII).[30] Блокирование этого притока или активации CaMKII предотвращает LTP, показывая, что это необходимые механизмы для LTP.[31] Кроме того, обилие CaMKII в синапсе вызывает LTP, показывая, что это причинный и достаточный механизм.[32]
CaMKII имеет несколько способов активации, чтобы вызвать включение рецепторов AMPA в перисинаптическую мембрану. Фермент CAMKII в конечном итоге отвечает за развитие актинового цитоскелета нейрональных клеток и, в конечном итоге, за развитие дендритов и аксонов (синаптическая пластичность).[33] Первый - прямое фосфорилирование синаптического белка 97 (SAP97 ).[34] Во-первых, SAP-97 и миозин-VI, моторный белок, связаны как комплекс с С-концом AMPAR. После фосфорилирования CaMKII комплекс перемещается в перисинаптическую мембрану.[35] Второй способ активации - через путь MAPK. CaMKII активирует белки Ras, которые затем активируют p42 / 44 MAPK, что приводит к встраиванию AMPAR непосредственно в перисинаптическую мембрану.[36]
Доставка рецептора AMPA к PSD в ответ на LTP
Как только рецепторы AMPA транспортируются в перисинаптическую область посредством фосфорилирования PKA или SAP97, рецепторы затем перемещаются в постсинаптическая плотность (PSD). Однако этот процесс передачи в PSD все еще остается спорным. Одна возможность состоит в том, что во время LTP происходит латеральное перемещение рецепторов AMPA от перисинпатических участков непосредственно к PSD.[37] Другая возможность состоит в том, что экзоцитоз внутриклеточных пузырьков напрямую отвечает за доставку AMPA в PSD.[38] Недавние данные свидетельствуют о том, что оба эти процесса происходят после стимула LTP; однако только латеральное движение рецепторов AMPA из перисинаптической области увеличивает количество рецепторов AMPA в PSD.[39] Точный механизм, ответственный за латеральное перемещение рецепторов AMPA к PSD, еще предстоит обнаружить; однако исследования открыли несколько важных белков для доставки рецепторов AMPA. Например, избыточная экспрессия SAP97 приводит к увеличению трафика рецептора AMPA в синапсы.[40] Было обнаружено, что помимо влияния на синаптическую локализацию, SAP97 влияет на проводимость рецептора AMPA в ответ на глутамат.[41] Миозин Белки представляют собой чувствительные к кальцию моторные белки, которые, как было обнаружено, также необходимы для доставки рецепторов AMPA. Нарушение взаимодействия миозина Vb с Rab11 и Rab11-FIP2 блокирует рост позвоночника и доставку рецептора AMPA.[42] Следовательно, возможно, что миозин может управлять боковым перемещением рецепторов AMPA в перисинпатической области к PSD. Трансмембранные регуляторные белки рецептора AMPA (TARP) представляют собой семейство белков, которые связываются с рецепторами AMPA и контролируют их транспортировку и проводимость.[43] CACNG2 (Старгазин) является одним из таких белков и, как обнаружено, связывает рецепторы AMPA в перисинаптических и постсинаптических областях.[44] Роль stargazin в перемещении между перисинаптическим и постсинаптическим регионами остается неясной; однако старгазин необходим для иммобилизации рецепторов AMPA в PSD путем взаимодействия с PSD-95.[45] PSD-95 стабилизирует рецепторы AMPA в синапсе, и нарушение взаимодействия звездгазин-PSD-95 подавляет синаптическую передачу.[46]
Конститутивная торговля и изменения в составе субъединиц
Рецепторы AMPA непрерывно доставляются (эндоцитируются, рециркулируются и повторно вводятся) в и из плазматическая мембрана. Переработка эндосом в дендритный позвоночник содержат пулы рецепторов AMPA для такой синаптической реинтеграции.[47] Существуют два различных пути доставки рецепторов AMPA: регулируемый путь и конститутивный путь.[48][49]
В регулируемом пути GluA1-содержащие рецепторы AMPA доставляются к синапсу зависимым от активности образом, стимулируемым Рецептор NMDA активация.[13] В базовых условиях регулируемый путь практически неактивен, временно активируясь только при индукции долгосрочное потенцирование.[47][48] Этот путь отвечает за укрепление синапсов и первоначальное формирование новых воспоминаний.[50]
В конститутивном пути рецепторы AMPA без GluA1, обычно гетеромерные рецепторы GluR2-GluR3, замещают рецепторы, содержащие GluA1, индивидуально, независимо от активности,[51][52] сохранение общего количества рецепторов AMPA в синапсе.[47][48] Этот путь отвечает за поддержание новых воспоминаний, поддерживая временные изменения, возникающие в результате регулируемого пути. В базовых условиях этот путь обычно активен, так как он необходим также для замещения поврежденных рецепторов.
Субъединицы GluA1 и GluA4 состоят из длинного карбокси (C) -хвоста, тогда как субъединицы GluA2 и GluA3 состоят из короткого карбокси-хвоста. Эти два пути регулируются взаимодействиями между С-концами субъединиц рецептора AMPA и синаптическими соединениями и белками. Длинные C-хвосты предотвращают вставку рецепторов GluR1 / 4 непосредственно в зону постсинаптической плотности (PSDZ) в отсутствие активности, тогда как короткие C-хвосты рецепторов GluA2 / 3 позволяют им вставляться непосредственно в PSDZ.[37][53] С-конец GluA2 взаимодействует и связывается с Чувствительный к N-этилмалеимиду гибридный белок,[54][55][56] что позволяет быстро вставлять GluR2-содержащие рецепторы AMPA в синапс.[57] Кроме того, субъединицы GluR2 / 3 более стабильно связаны с синапсом, чем субъединицы GluR1.[58][59][60]
LTD-индуцированный эндоцитоз рецепторов AMPA
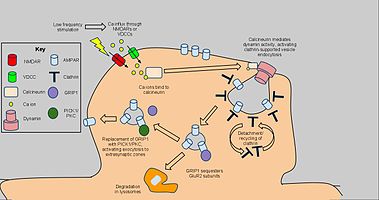
Длительная депрессия задействует механизмы снижения плотности рецепторов AMPA в выбранных дендритных шипах, в зависимости от клатрин и кальциневрин и в отличие от конститутивного трафика AMPAR. Стартовый сигнал для AMPAR эндоцитоз представляет собой NMDAR-зависимый приток кальция от низкочастотной стимуляции, который, в свою очередь, активирует протеинфосфатазы PP1 и кальциневрин. Однако эндоцитоз AMPAR также активируется потенциал-зависимые кальциевые каналы, агонизм рецепторов AMPA и введение инсулин, предполагая общий приток кальция как причину эндоцитоза AMPAR.[61] Блокирование PP1 не предотвращало эндоцитоз AMPAR, но применение антагониста кальциневрина приводило к значительному ингибированию этого процесса.[62]
Кальциневрин взаимодействует с эндоцитотическим комплексом в постсинаптической зоне, объясняя его влияние на LTD.[63] Комплекс, состоящий из покрытой клатрином ямки под частью AMPAR-содержащей плазматической мембраны и взаимодействующих белков, является прямым механизмом восстановления AMPAR, в частности рецепторов, содержащих субъединицу GluR2 / GluR3, в синапсе. Взаимодействие с кальциневрином активирует динамин Активность ГТФазы, позволяющая клатриновой ямке вырезать себя из клеточной мембраны и становиться цитоплазматическим пузырьком.[64] После отделения клатриновой оболочки другие белки могут напрямую взаимодействовать с AMPAR, используя ПДЗ карбоксильные хвостовые домены; например, белок 1, взаимодействующий с глутаматным рецептором (GRIP1 ) участвует во внутриклеточной секвестрации AMPAR.[65] Затем внутриклеточные AMPAR сортируются на предмет деградации лизосомами или рециркуляции в клеточную мембрану.[66] Для последнего PICK1 и PKC может вытеснять GRIP1, чтобы вернуть AMPAR на поверхность, обращая вспять эффекты эндоцитоза и LTD. при необходимости.[67] Тем не менее, упомянутый выше кальций-зависимый, опосредованный динамином механизм был задействован как ключевой компонент LTD. и как таковые могут иметь приложения для дальнейших поведенческих исследований.[68]
Роль в припадках
Рецепторы AMPA играют ключевую роль в возникновении и распространении эпилептических припадков.[69] Каиновая кислота, судорожное средство, которое широко используется в исследованиях эпилепсии, вызывает судороги, частично за счет активации рецепторов AMPA.[70]
Молекулярная мишень для терапии эпилепсии
Неконкурентные антагонисты рецепторов AMPA талампанель и перампанель продемонстрировали свою активность при лечении парциальных припадков у взрослых,[71][72] это указывает на то, что антагонисты рецептора AMPA представляют собой потенциальную мишень для лечения эпилепсии.[73][74] Перампанель (торговое название: Fycompa) получил одобрение Европейской Комиссией на получение регистрационного удостоверения для лечения частичной эпилепсии 27 июля 2012 г. Препарат был одобрен в Соединенных Штатах Управление по контролю за продуктами и лекарствами (FDA) 22 октября 2012 г. Как и в случае последних разработанных ПЭП, включая прегабалин, лакозамид и эзогабин, FDA рекомендовало классифицировать перампанель по отдел по борьбе с наркотиками (DEA) как контролируемый препарат. Он был обозначен как контролируемое вещество Списка 3.
Декановая кислота действует как неконкурентный антагонист рецептора AMPA в терапевтически релевантных концентрациях в зависимости от напряжения и субъединицы, и этого достаточно, чтобы объяснить его противосудорожные эффекты.[75] Это прямое ингибирование возбуждающей нейротрансмиссии декановой кислотой в головном мозге способствует противосудорожному эффекту триглицерид со средней длиной цепи кетогенная диета.[75] Декановая кислота и антагонист рецептора AMPA препарат перампанел действуют на разных участках рецептора AMPA, и поэтому возможно, что они оказывают совместное действие на рецептор AMPA, предполагая, что перампанел и кетогенная диета могут быть синергическими.[75]
Доклинические исследования показывают, что несколько производных ароматических аминокислот с антиглутаматергическими свойствами, включая антагонизм рецептора AMPA и ингибирование высвобождения глутамата, такие как 3,5-дибром-D-тирозин и 3,5-дибром-L-фенилалнин, проявляют сильный противосудорожный эффект на животных моделях. предполагая использование этих соединений в качестве нового класса противоэпилептических препаратов.[76][77]
Агонисты

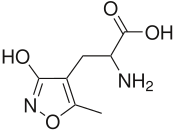
- 5-фторовиллардиин - синтетическая модификация Willardiine
- AMPA - синтетический агонист, в честь которого назван рецептор
- Домоевая кислота - агонист природного происхождения, вызывающий амнестическое отравление моллюсками
- Глютаминовая кислота (глутамат) - эндогенный агонист
- Иботеновая кислота - агонист природного происхождения, обнаруженный в Мухомор мухомор
- Квискваловая кислота - естественный агонист, обнаруженный у определенных видов
- Willardiine - агонист природного происхождения
Положительные аллостерические модуляторы
- Анирацетам
- Циклотиазид
- CX-516
- CX-546
- CX-614
- CX-717
- Фарампатор (CX-691, ORG-24448)
- IDRA-21
- LY-404187
- LY-503430[78][79]
- Мибампатор (LY-451395)
- ORG-26576
- Оксирацетам
- PEPA
- PF-04958242
- Пирацетам
- Прамирацетам
- Tulrampator (S-47445, CX-1632)
Антагонисты
- Бекампанель
- CNQX
- Дасолампанель
- DNQX
- Фанапанель (MPQX)
- Кайтоцефалин
- Кинуреновая кислота - эндогенный лиганд
- L-теанин
- NBQX
- 3,5-Дибром-L-фенилаланин, встречающееся в природе галогенированное производное L-фенилаланин[80]
- Селурампанель
- Тезампанель
- Zonampanel
Отрицательные аллостерические модуляторы

- Барбитураты (например., пентобарбитал, тиопентал натрия ) - неселективный
- Этиловый спирт - неизбирательный
- Ингаляционные анестетики (например., циклопропан, энфлуран, галотан, изофлуран, севофлуран ) - неселективный
- GYKI-52466
- Ирампанель
- Перампанель
- Талампанель
- PEP1-TGL: аналог пептида С-конца субъединицы GluA1, который ингибирует включение рецептора AMPA в постсинаптическую плотность[81][82]
Смотрите также
использованная литература
- ^ Оноре Т., Лауридсен Дж, Крогсгаард-Ларсен П. (январь 1982 г.). «Связывание [3H] AMPA, структурного аналога глутаминовой кислоты, с мембранами головного мозга крысы». Журнал нейрохимии. 38 (1): 173–8. Дои:10.1111 / j.1471-4159.1982.tb10868.x. PMID 6125564.
- ^ Армстронг Н. (2000). "Армстронг и Гуо (2000) Механизмы активации и антагонизма AMPA-чувствительного рецептора глутамата: кристаллические структуры ядра связывания лиганда GluR2". Нейрон. 28 (1): 165–181. Дои:10.1016 / S0896-6273 (00) 00094-5. PMID 11086992. S2CID 3128719.
- ^ "Рецепторы глутамата: структуры и функции. Центр синаптической пластичности Бристольского университета". Архивировано из оригинал 15 сентября 2007 г.. Получено 2007-09-02.
- ^ Ши С.Х., Хаяси Ю., Петралия Р.С., Заман С.Х., Вентхольд Р.Дж., Свобода К., Малинов Р. (июнь 1999 г.). «Быстрая доставка по позвоночнику и перераспределение рецепторов AMPA после активации синаптического рецептора NMDA». Наука. 284 (5421): 1811–6. CiteSeerX 10.1.1.376.3281. Дои:10.1126 / science.284.5421.1811. PMID 10364548.
- ^ Песня I, Хуганир Р.Л. (ноябрь 2002 г.). «Регулирование рецепторов AMPA при синаптической пластичности». Тенденции в неврологии. 25 (11): 578–88. Дои:10.1016 / S0166-2236 (02) 02270-1. PMID 12392933. S2CID 1993509.
- ^ а б Майер М.Л. (июнь 2005 г.). «Ионные каналы глутаматного рецептора» (PDF). Текущее мнение в нейробиологии. 15 (3): 282–8. Дои:10.1016 / j.conb.2005.05.004. PMID 15919192. S2CID 39812856.
- ^ а б Грегер И. Х., Зифф Е.Б., Пенн А.С. (август 2007 г.). «Молекулярные детерминанты сборки субъединицы рецептора AMPA». Тенденции в неврологии. 30 (8): 407–16. Дои:10.1016 / j.tins.2007.06.005. PMID 17629578. S2CID 7505830.
- ^ Холлманн М., Марон С., Хайнеманн С. (декабрь 1994 г.). «Маркировка сайта N-гликозилирования предполагает топологию трех трансмембранных доменов для рецептора глутамата GluR1». Нейрон. 13 (6): 1331–43. Дои:10.1016/0896-6273(94)90419-7. PMID 7993626. S2CID 39682094.
- ^ Леонард А.С., Даваре М.А., Хорн М.С., Гарнер СС, Ад JW (июль 1998 г.). «SAP97 связан с субъединицей GluR1 рецептора альфа-амино-3-гидрокси-5-метилизоксазол-4-пропионовой кислоты». Журнал биологической химии. 273 (31): 19518–24. Дои:10.1074 / jbc.273.31.19518. PMID 9677374.
- ^ а б Грегер И.Х., Хатри Л., Зифф Е.Б. (май 2002 г.). «Редактирование РНК на arg607 контролирует выход рецептора AMPA из эндоплазматического ретикулума». Нейрон. 34 (5): 759–72. Дои:10.1016 / S0896-6273 (02) 00693-1. PMID 12062022. S2CID 15936250.
- ^ Летучие мыши C, Groc L, Choquet D (март 2007 г.). «Взаимодействие между Stargazin и PSD-95 регулирует транспортировку через поверхность рецептора AMPA». Нейрон. 53 (5): 719–34. Дои:10.1016 / j.neuron.2007.01.030. PMID 17329211. S2CID 16423733.
- ^ Бём Дж., Канг М.Г., Джонсон Р.К., Эстебан Дж., Хуганир Р.Л., Малинов Р. (июль 2006 г.). «Синаптическое включение рецепторов AMPA во время LTP контролируется сайтом фосфорилирования PKC на GluR1». Нейрон. 51 (2): 213–25. Дои:10.1016 / j.neuron.2006.06.013. PMID 16846856. S2CID 16208091.
- ^ а б Hayashi Y, Shi SH, Esteban JA, Piccini A, Poncer JC, Malinow R (март 2000 г.). «Управление рецепторами AMPA в синапсах с помощью LTP и CaMKII: требование для взаимодействия GluR1 и домена PDZ». Наука. 287 (5461): 2262–7. Дои:10.1126 / science.287.5461.2262. PMID 10731148. S2CID 17001488.
- ^ Деркач В., Баррия А., Содерлинг Т. Р. (март 1999 г.). «Са2 + / кальмодулин-киназа II увеличивает проводимость канала глутаматных рецепторов альфа-амино-3-гидрокси-5-метил-4-изоксазолепропионата». Труды Национальной академии наук Соединенных Штатов Америки. 96 (6): 3269–74. Дои:10.1073 / пнас.96.6.3269. ЧВК 15931. PMID 10077673.
- ^ Delgado JY, Coba M, Anderson CN, Thompson KR, Gray EE, Heusner CL, et al. (Ноябрь 2007 г.). «Активация рецептора NMDA дефосфорилирует субъединицы рецептора глутамата 1 рецептора AMPA по треонину 840». Журнал неврологии. 27 (48): 13210–21. Дои:10.1523 / JNEUROSCI.3056-07.2007. ЧВК 2851143. PMID 18045915.
- ^ Банке Т.Г., Боуи Д., Ли Х., Хуганир Р.Л., Шоусбой А., Трайнелис С.Ф. (январь 2000 г.). «Контроль функции рецептора GluR1 AMPA с помощью цАМФ-зависимой протеинкиназы». Журнал неврологии. 20 (1): 89–102. Дои:10.1523 / JNEUROSCI.20-01-00089.2000. ЧВК 6774102. PMID 10627585.
- ^ Армстронг Н., Сунь И, Чен Г.К., Гуо Э. (октябрь 1998 г.). «Структура ядра, связывающего лиганд глутаматного рецептора в комплексе с каинатом». Природа. 395 (6705): 913–7. Дои:10.1038/27692. PMID 9804426. S2CID 4405926.
- ^ а б Platt SR (март 2007 г.). «Роль глутамата в здоровье и болезнях центральной нервной системы - обзор». Ветеринарный журнал. 173 (2): 278–86. Дои:10.1016 / j.tvjl.2005.11.007. PMID 16376594.
- ^ Розенмунд С., Стерн-Бах И., Стивенс К.Ф. (июнь 1998 г.). «Тетрамерная структура канала рецептора глутамата». Наука. 280 (5369): 1596–9. Дои:10.1126 / science.280.5369.1596. HDL:11858 / 00-001M-0000-0012-FDD8-B. PMID 9616121.
- ^ Армстронг Н., Джасти Дж., Бейх-Франдсен М., Гуо Э. (октябрь 2006 г.). «Измерение конформационных изменений, сопровождающих десенсибилизацию в ионотропном рецепторе глутамата». Ячейка. 127 (1): 85–97. Дои:10.1016 / j.cell.2006.08.037. PMID 17018279. S2CID 16564029.
- ^ Kim DY, Kim SH, Choi HB, Min C, Gwag BJ (июнь 2001 г.). «Высокое содержание мРНК GluR1 и уменьшенное редактирование Q / R мРНК GluR2 в отдельных нейронах NADPH-диафоразы». Молекулярная и клеточная нейронауки. 17 (6): 1025–33. Дои:10.1006 / mcne.2001.0988. PMID 11414791. S2CID 15351461.
- ^ Люшер С., Маленка RC (февраль 2011 г.). «Синаптическая пластичность, вызванная лекарствами при зависимости: от молекулярных изменений до ремоделирования цепей». Нейрон. 69 (4): 650–63. Дои:10.1016 / j.neuron.2011.01.017. ЧВК 4046255. PMID 21338877.
- ^ Мосбахер Дж., Шопфер Р., Моньер Х., Бурнашев Н., Зеебург PH, Рупперсберг Дж. П. (ноябрь 1994 г.). «Молекулярный детерминант субмиллисекундной десенсибилизации в рецепторах глутамата». Наука. 266 (5187): 1059–62. Дои:10.1126 / science.7973663. PMID 7973663.
- ^ Соммер Б., Кейнянен К., Вердорн Т.А., Висден В., Бурнашев Н., Херб А. и др. (Сентябрь 1990 г.). «Flip and flop: клеточно-специфический функциональный переключатель в глутаматных каналах ЦНС». Наука. 249 (4976): 1580–5. Дои:10.1126 / science.1699275. PMID 1699275.
- ^ Пей В., Хуанг З., Ню Л. (февраль 2007 г.). «GluR3 flip and flop: различия в кинетике открытия каналов». Биохимия. 46 (7): 2027–36. Дои:10.1021 / bi062213s. PMID 17256974.
- ^ Иствуд С.Л., Бернет П.В., Харрисон П.Дж. (февраль 1997 г.). «Изоформы flip и flop субъединицы рецептора глутамата GluR2 уменьшаются в формировании гиппокампа при шизофрении: исследование полимеразной цепной реакции с обратной транскриптазой (ОТ-ПЦР)». Исследование мозга. Молекулярные исследования мозга. 44 (1): 92–8. Дои:10.1016 / s0169-328x (96) 00195-7. PMID 9030702.
- ^ Whitlock JR, Heynen AJ, Shuler MG, Bear MF (август 2006 г.). «Обучение вызывает долгосрочное потенцирование в гиппокампе». Наука. 313 (5790): 1093–7. Дои:10.1126 / science.1128134. PMID 16931756. S2CID 612352.
- ^ Марен С., Токко Дж., Стэндли С., Бодри М., Томпсон РФ (октябрь 1993 г.). «Постсинаптические факторы в экспрессии долгосрочной потенциации (LTP): усиление связывания рецептора глутамата после индукции LTP in vivo». Труды Национальной академии наук Соединенных Штатов Америки. 90 (20): 9654–8. Дои:10.1073 / пнас.90.20.9654. ЧВК 47628. PMID 8415757.
- ^ Перкинтон М.С., Сихра Т.С., Уильямс Р.Дж. (июль 1999 г.). «Са (2 +) - проницаемые рецепторы AMPA индуцируют фосфорилирование белка, связывающего элемент ответа цАМФ, посредством зависимой от фосфатидилинозитол 3-киназы стимуляции сигнального каскада митоген-активируемой протеинкиназы в нейронах». Журнал неврологии. 19 (14): 5861–74. Дои:10.1523 / JNEUROSCI.19-14-05861.1999. ЧВК 6783096. PMID 10407026.
- ^ Фукунага К., Стоппини Л., Миямото Е., Мюллер Д. (апрель 1993 г.). «Долгосрочное усиление связано с повышенной активностью Са2 + / кальмодулин-зависимой протеинкиназы II». Журнал биологической химии. 268 (11): 7863–7. PMID 8385124.
- ^ Лисман Дж, Шульман Х, Клайн Х (март 2002 г.). «Молекулярная основа функции CaMKII в синаптической и поведенческой памяти». Обзоры природы. Неврология. 3 (3): 175–90. Дои:10.1038 / номер 753. PMID 11994750. S2CID 5844720.
- ^ Маммен А.Л., Камеяма К., Рош К.В., Хуганир Р.Л. (декабрь 1997 г.). «Фосфорилирование субъединицы рецептора GluR1 альфа-амино-3-гидрокси-5-метилизоксазол-4-пропионовой кислоты кальций / кальмодулин-зависимой киназой II». Журнал биологической химии. 272 (51): 32528–33. Дои:10.1074 / jbc.272.51.32528. PMID 9405465.
- ^ Эберт Д.Х., Гринберг МЭ (январь 2013 г.). «Активно-зависимая нейрональная сигнализация и расстройство аутистического спектра». Природа. 493 (7432): 327–37. Дои:10.1038 / природа11860. ЧВК 3576027. PMID 23325215.
- ^ Маусери Д., Каттабени Ф., Ди Лука М., Гардони Ф. (май 2004 г.). «Фосфорилирование кальций / кальмодулин-зависимой протеинкиназы II направляет связанный с синапсом белок 97 в шипы». Журнал биологической химии. 279 (22): 23813–21. Дои:10.1074 / jbc.M402796200. PMID 15044483.
- ^ Ву Х, Нэш Дж. Э., Заморано П., Гарнер СС (август 2002 г.). «Взаимодействие SAP97 с актиновым моторным миозином VI, направленным на минус-конец. Последствия для передачи рецепторов AMPA». Журнал биологической химии. 277 (34): 30928–34. Дои:10.1074 / jbc.M203735200. PMID 12050163.
- ^ Чжу Дж.Дж., Цинь Ю., Чжао М., Ван Элст Л., Малинов Р. (август 2002 г.). «Ras и Rap контролируют движение рецепторов AMPA во время синаптической пластичности». Ячейка. 110 (4): 443–55. Дои:10.1016 / S0092-8674 (02) 00897-8. PMID 12202034. S2CID 12858091.
- ^ а б Borgdorff AJ, Choquet D (июнь 2002 г.). «Регуляция боковых движений рецепторов AMPA». Природа. 417 (6889): 649–53. Дои:10.1038 / природа00780. PMID 12050666. S2CID 4422115.
- ^ Park M, Penick EC, Edwards JG, Kauer JA, Ehlers MD (сентябрь 2004 г.). «Вторичные эндосомы поставляют рецепторы AMPA для LTP». Наука. 305 (5692): 1972–5. Дои:10.1126 / science.1102026. PMID 15448273. S2CID 34651431.
- ^ Макино Х., Малинов Р. (ноябрь 2009 г.). «Включение рецептора AMPA в синапсы во время LTP: роль бокового движения и экзоцитоза». Нейрон. 64 (3): 381–90. Дои:10.1016 / j.neuron.2009.08.035. ЧВК 2999463. PMID 19914186.
- ^ Ховард М.А., Элиас Г.М., Элиас Л.А., Сват В., Николл Р.А. (февраль 2010 г.). «Роль SAP97 в динамике синаптических рецепторов глутамата». Труды Национальной академии наук Соединенных Штатов Америки. 107 (8): 3805–10. Дои:10.1073 / pnas.0914422107. ЧВК 2840522. PMID 20133708.
- ^ Waites CL, Specht CG, Härtel K, Leal-Ortiz S, Genoux D, Li D и др. (Апрель 2009 г.). «Изоформы синаптического SAP97 регулируют динамику рецептора AMPA и доступ к пресинаптическому глутамату». Журнал неврологии. 29 (14): 4332–45. Дои:10.1523 / JNEUROSCI.4431-08.2009. ЧВК 3230533. PMID 19357261.
- ^ Ван З., Эдвардс Дж. Г., Райли Н., Прованс Д. В., Карчер Р., Ли XD и др. (Октябрь 2008 г.). «Миозин Vb мобилизует рециклинг эндосом и рецепторов AMPA для постсинаптической пластичности». Ячейка. 135 (3): 535–48. Дои:10.1016 / j.cell.2008.09.057. ЧВК 2585749. PMID 18984164.
- ^ Николл Р.А., Томита С., Бредт Д.С. (март 2006 г.). «Вспомогательные субъединицы помогают рецепторам глутамата AMPA-типа». Наука. 311 (5765): 1253–6. Дои:10.1126 / science.1123339. PMID 16513974. S2CID 40782882.
- ^ Томита С., Чен Л., Кавасаки Ю., Петралиа Р.С., Вентхольд Р.Дж., Николл Р.А., Бредт Д.С. (май 2003 г.). «Функциональные исследования и распространение определяют семейство белков, регулирующих трансмембранный рецептор AMPA». Журнал клеточной биологии. 161 (4): 805–16. Дои:10.1083 / jcb.200212116. ЧВК 2199354. PMID 12771129.
- ^ Чен Л., Четкович Д.М., Петралиа Р.С., Суини Н.Т., Кавасаки Ю., Вентхольд Р.Дж. и др. (2000). «Старгазин регулирует синаптическое нацеливание на рецепторы AMPA с помощью двух различных механизмов». Природа. 408 (6815): 936–43. Дои:10.1038/35050030. PMID 11140673. S2CID 4427689.
- ^ Летучие мыши C, Groc L, Choquet D (март 2007 г.). «Взаимодействие между Stargazin и PSD-95 регулирует транспортировку через поверхность рецептора AMPA». Нейрон. 53 (5): 719–34. Дои:10.1016 / j.neuron.2007.01.030. PMID 17329211. S2CID 16423733.
- ^ а б c Пастух JD, Хуганир RL (2007). «Клеточная биология синаптической пластичности: трафик рецепторов AMPA». Ежегодный обзор клеточной биологии и биологии развития. 23: 613–43. Дои:10.1146 / annurev.cellbio.23.090506.123516. PMID 17506699. S2CID 7048661.
- ^ а б c Малинов Р., Майнен З.Ф., Хаяши Ю. (июнь 2000 г.). «Механизмы LTP: от тишины к четырехполосному движению». Текущее мнение в нейробиологии. 10 (3): 352–7. Дои:10.1016 / S0959-4388 (00) 00099-4. PMID 10851179. S2CID 511079.
- ^ Маленка RC (ноябрь 2003 г.). «Синаптическая пластичность и трафик рецепторов AMPA». Летопись Нью-Йоркской академии наук. 1003: 1–11. Дои:10.1196 / анналы.1300.001. PMID 14684431.
- ^ Кесселс Х.В., Малинов Р. (февраль 2009 г.). «Пластичность и поведение синаптических AMPA-рецепторов». Нейрон. 61 (3): 340–50. Дои:10.1016 / j.neuron.2009.01.015. ЧВК 3917551. PMID 19217372.
- ^ Маккормак С.Г., Сторнетта Р.Л., Чжу Дж.Дж. (апрель 2006 г.). «Обмен синаптического рецептора AMPA поддерживает двунаправленную пластичность». Нейрон. 50 (1): 75–88. Дои:10.1016 / j.neuron.2006.02.027. PMID 16600857. S2CID 17478776.
- ^ Чжу Дж. Дж., Эстебан Дж. А., Хаяши Ю., Малинов Р. (ноябрь 2000 г.). «Постнатальная синаптическая потенциация: доставка GluR4-содержащих рецепторов AMPA путем спонтанной активности». Природа Неврология. 3 (11): 1098–106. Дои:10.1038/80614. HDL:10261/47079. PMID 11036266. S2CID 16116261.
- ^ Passafaro M, Piëch V, Sheng M (сентябрь 2001 г.). «Субъединично-специфические временные и пространственные паттерны экзоцитоза рецептора AMPA в нейронах гиппокампа». Природа Неврология. 4 (9): 917–26. Дои:10.1038 / nn0901-917. PMID 11528423. S2CID 32852272.
- ^ Песня I, Камбодж С., Ся Дж, Донг Х, Ляо Д., Хуганир Р.Л. (август 1998 г.). «Взаимодействие фактора, чувствительного к N-этилмалеимиду, с рецепторами AMPA». Нейрон. 21 (2): 393–400. Дои:10.1016 / S0896-6273 (00) 80548-6. PMID 9728920.
- ^ Остен П., Шривастава С., Инман Г.Дж., Вилим Ф.С., Хатри Л., Ли Л.М. и др. (Июль 1998 г.). «С-конец GluR2 рецептора AMPA может опосредовать обратимое, АТФ-зависимое взаимодействие с NSF и альфа- и бета-SNAP». Нейрон. 21 (1): 99–110. Дои:10.1016 / S0896-6273 (00) 80518-8. PMID 9697855. S2CID 18569829.
- ^ Нишимун А., Исаак Дж. Т., Мольнар Э., Ноэль Дж., Нэш С. Р., Тагая М. и др. (Июль 1998 г.). «Связывание NSF с GluR2 регулирует синаптическую передачу». Нейрон. 21 (1): 87–97. Дои:10.1016 / S0896-6273 (00) 80517-6. HDL:2433/180867. PMID 9697854. S2CID 18956893.
- ^ Beretta F, Sala C, Saglietti L, Hirling H, Sheng M, Passafaro M (апрель 2005 г.). «Взаимодействие с NSF важно для прямой вставки GluR2 в синаптические сайты». Молекулярная и клеточная нейронауки. 28 (4): 650–60. Дои:10.1016 / j.mcn.2004.11.008. PMID 15797712. S2CID 46716417.
- ^ Cingolani LA, Thalhammer A, Yu LM, Catalano M, Ramos T., Colicos MA, Goda Y (июнь 2008 г.). «Зависимая от активности регуляция состава и количества синаптических рецепторов AMPA с помощью интегринов бета3». Нейрон. 58 (5): 749–62. Дои:10.1016 / j.neuron.2008.04.011. ЧВК 2446609. PMID 18549786.
- ^ Saglietti L, Dequidt C, Kamieniarz K, Rousset MC, Valnegri P, Thoumine O и др. (Май 2007 г.). «Внеклеточные взаимодействия между GluR2 и N-кадгерином в регуляции позвоночника». Нейрон. 54 (3): 461–77. Дои:10.1016 / j.neuron.2007.04.012. PMID 17481398. S2CID 14600986.
- ^ Сильверман Дж. Б., Реституито С., Лу В., Ли-Эдвардс Л., Хатри Л., Зифф Е.Б. (август 2007 г.). «Синаптическое закрепление рецепторов AMPA кадгеринами через нейральный белок руки, связанный с плакофилином, белковые комплексы, связывающие рецептор AMPA». Журнал неврологии. 27 (32): 8505–16. Дои:10.1523 / JNEUROSCI.1395-07.2007. ЧВК 6672939. PMID 17687028.
- ^ Кэрролл Р.К., Битти Е.К., Ся Х, Люшер С., Альтшулер Ю., Николл Р.А. и др. (Ноябрь 1999 г.). «Динамин-зависимый эндоцитоз ионотропных глутаматных рецепторов». Труды Национальной академии наук Соединенных Штатов Америки. 96 (24): 14112–7. Дои:10.1073 / пнас.96.24.14112. ЧВК 24199. PMID 10570207.
- ^ Beattie EC, Carroll RC, Yu X, Morishita W, Yasuda H, von Zastrow M, Malenka RC (декабрь 2000 г.). «Регулирование эндоцитоза рецептора AMPA с помощью механизма передачи сигналов, общего с LTD». Природа Неврология. 3 (12): 1291–300. Дои:10.1038/81823. PMID 11100150.
- ^ Лай М.М., Хонг Дж. Дж., Руджеро А.М., Бернетт П.Е., Слепнев В.И., Де Камилли П., Снайдер С.Х. (сентябрь 1999 г.). «Комплекс кальциневрин-динамин 1 как сенсор кальция для эндоцитоза синаптических везикул». Журнал биологической химии. 274 (37): 25963–6. Дои:10.1074 / jbc.274.37.25963. PMID 10473536.
- ^ Юнг Н., Хаук В. (сентябрь 2007 г.). «Клатрин-опосредованный эндоцитоз в синапсах». Движение. 8 (9): 1129–36. Дои:10.1111 / j.1600-0854.2007.00595.x. PMID 17547698.
- ^ Доу М.И., Читтаджаллу Р., Бортолотто З.А., Дев К.К., Дюпрат Ф., Хенли Дж.М. и др. (Декабрь 2000 г.). «Белки PDZ, взаимодействующие с C-концевым GluR2 / 3, участвуют в PKC-зависимой регуляции рецепторов AMPA в синапсах гиппокампа». Нейрон. 28 (3): 873–86. Дои:10.1016 / S0896-6273 (00) 00160-4. HDL:2262/89240. PMID 11163273. S2CID 13727678.
- ^ Ehlers MD (ноябрь 2000 г.). «Повторное внедрение или деградация рецепторов AMPA, определяемая зависимым от активности эндоцитарным сортированием». Нейрон. 28 (2): 511–25. Дои:10.1016 / S0896-6273 (00) 00129-X. PMID 11144360. S2CID 16333109.
- ^ Лу В., Зифф Е.Б. (август 2005 г.). «PICK1 взаимодействует с ABP / GRIP для регулирования передачи рецептора AMPA». Нейрон. 47 (3): 407–21. Дои:10.1016 / j.neuron.2005.07.006. PMID 16055064. S2CID 17100359.
- ^ Ван Ю.Т. (март 2008 г.). «Исследование роли эндоцитоза AMPAR и длительной депрессии в поведенческой сенсибилизации: актуальность для лечения заболеваний головного мозга, включая наркозависимость». Британский журнал фармакологии. 153 Приложение 1 (S1): S389-95. Дои:10.1038 / sj.bjp.0707616. ЧВК 2268058. PMID 18059315.
- ^ Рогавский М.А. (2013). «Рецепторы AMPA как молекулярная мишень в терапии эпилепсии». Acta Neurologica Scandinavica. Дополнение. 127 (197): 9–18. Дои:10.1111 / ane.12099. ЧВК 4506648. PMID 23480151.
- ^ Фрич Б., Рейс Дж., Гасиор М., Камински Р.М., Рогавски М.А. (апрель 2014 г.). «Роль каинатных рецепторов GluK1 в припадках, эпилептических разрядах и эпилептогенезе». Журнал неврологии. 34 (17): 5765–75. Дои:10.1523 / JNEUROSCI.5307-13.2014. ЧВК 3996208. PMID 24760837.
- ^ Биалер М., Йоханнесен С.И., Купферберг Х.Дж., Леви Р.Х., Перука Э., Томсон Т. (январь 2007 г.). «Отчет о ходе разработки новых противоэпилептических препаратов: резюме Восьмой Эйлатской конференции (EILAT VIII)». Исследования эпилепсии. 73 (1): 1–52. Дои:10.1016 / j.eplepsyres.2006.10.008. PMID 17158031. S2CID 45026113.
- ^ French JA, Krauss GL, Biton V, Squillacote D, Yang H, Laurenza A и др. (Август 2012 г.). «Дополнительный перампанел при рефрактерных приступах с частичным началом: рандомизированное исследование III фазы 304». Неврология. 79 (6): 589–96. Дои:10.1212 / WNL.0b013e3182635735. ЧВК 3413761. PMID 22843280.
- ^ Рогавский М.А. (март 2011 г.). «Возвращение к рецепторам AMPA как мишени противоэпилептического препарата». Течение эпилепсии. 11 (2): 56–63. Дои:10.5698/1535-7511-11.2.56. ЧВК 3117497. PMID 21686307.
- ^ Сакаи Ф, Игараси Х, Судзуки С, Тадзаки Й (1989). «Церебральный кровоток и церебральный гематокрит у пациентов с церебральной ишемией, измеренные с помощью однофотонной эмиссионной компьютерной томографии». Acta Neurologica Scandinavica. Дополнение. 127: 9–13. Дои:10.1111 / j.1600-0404.1989.tb01805.x. PMID 2631521. S2CID 30934688.
- ^ а б c Чанг П., Огюстен К., Боддум К., Уильямс С., Сан М., Тершак Дж. А. и др. (Февраль 2016). «Контроль приступов декановой кислотой посредством прямого ингибирования рецепторов AMPA». Мозг. 139 (Pt 2): 431–43. Дои:10.1093 / мозг / awv325. ЧВК 4805082. PMID 26608744.
- ^ Цао В., Шах Х.П., Глушаков А.В., Мекка А.П., Ши П., Самнерс С. и др. (Декабрь 2009 г.). «Эффективность 3,5-дибром-L-фенилаланина в моделях инсульта, судорог и сенсомоторного стробирования у крыс». Британский журнал фармакологии. 158 (8): 2005–13. Дои:10.1111 / j.1476-5381.2009.00498.x. ЧВК 2807662. PMID 20050189.
- ^ Цао В., Глушаков А., Шах Х. П., Мекка А. П., Самнерс С., Ши П. и др. (Апрель 2011 г.). «Галогенированная ароматическая аминокислота 3,5-дибром-D: -тирозин оказывает благотворное влияние при экспериментальном инсульте и судорогах». Аминокислоты. 40 (4): 1151–8. Дои:10.1007 / s00726-010-0739-4. PMID 20839013. S2CID 19852158.
- ^ Мюррей Т.К., Уолли К., Робинсон С.С., Уорд М.А., Хикс Калифорния, Лодж Д. и др. (Август 2003 г.). «LY503430, новый усилитель рецептора альфа-амино-3-гидрокси-5-метилизоксазол-4-пропионовой кислоты с функциональным, нейропротекторным и нейротрофическим действием в моделях болезни Паркинсона на грызунах». Журнал фармакологии и экспериментальной терапии. 306 (2): 752–62. Дои:10.1124 / jpet.103.049445. PMID 12730350. S2CID 86751458.
- ^ О'Нил М.Дж., Бликман Д., Циммерман Д.М., Нисенбаум Е.С. (июнь 2004 г.). «Потенциаторы рецепторов AMPA для лечения расстройств ЦНС». Текущие целевые показатели по лекарствам. ЦНС и неврологические расстройства. 3 (3): 181–94. Дои:10.2174/1568007043337508. PMID 15180479.
- ^ Яроцкий В., Глушаков А.В., Самнерс С., Гравенштейн Н., Деннис Д.М., Зеуберт С.Н., Мартынюк А.Е. (май 2005 г.). «Дифференциальная модуляция глутаматергической передачи 3,5-дибром-L-фенилаланином». Молекулярная фармакология. 67 (5): 1648–54. Дои:10.1124 / моль 104.005983. PMID 15687225. S2CID 11672391.
- ^ "Hayashi et al. (200) Включение рецепторов AMPA в синапсы с помощью LTP и CaMKII: требование для взаимодействия GluR1 и PDZ-доменов. Science 287; 2262-2267".
- ^ Tazerart S, Mitchell DE, Miranda-Rottmann S, Araya R (август 2020 г.). «Правило пластичности дендритных шипов, зависящее от времени спайков». Nature Communications. 11 (1): 4276. Дои:10.1038 / s41467-020-17861-7. ЧВК 7449969. PMID 32848151.