Никотиновый рецептор ацетилхолина - Nicotinic acetylcholine receptor


Никотиновые рецепторы ацетилхолина, или же нАХР, находятся рецептор полипептиды которые отвечают на нейротрансмиттер ацетилхолин. Никотиновые рецепторы также реагируют на такие препараты, как агонист. никотин. Они обнаруживаются в центральной и периферической нервной системе, мышцах и многих других тканях многих организмов. На нервномышечное соединение они являются первичными рецепторами в мышцах для связи двигательных нервов и мышц, которые контролируют сокращение мышц. В периферической нервной системе: (1) они передают исходящие сигналы от пресинаптических к постсинаптическим клеткам симпатической и парасимпатической нервной системы, и (2) они являются рецепторами, обнаруженными на скелетных мышцах, которые получают ацетилхолин, высвобождаемый для сигнала мышечного сокращения. В иммунной системе nAChR регулируют воспалительные процессы и передают сигнал через различные внутриклеточные пути.[1] В насекомые, то холинергический система ограничена Центральная нервная система.[2]
Никотиновые рецепторы считаются холинэргические рецепторы, поскольку они реагируют на ацетилхолин. Никотиновые рецепторы получили свое название от никотин который не стимулирует мускариновые рецепторы ацетилхолина но вместо этого избирательно связывается с никотиновыми рецепторами.[3][4][5] Мускариновый рецептор ацетилхолина также получил свое название от химического вещества, которое избирательно прикрепляется к этому рецептору - мускарин.[6] Сам по себе ацетилхолин связывается как с мускариновыми, так и с никотиновыми рецепторами ацетилхолина.[7]
В качестве ионотропный рецепторы, nAChR напрямую связаны с ионными каналами. Новые данные свидетельствуют о том, что эти рецепторы также могут использовать вторые мессенджеры (в качестве метаботропные рецепторы делать) в некоторых случаях.[8] Никотиновые ацетилхолиновые рецепторы являются наиболее изученными из ионотропных рецепторов.[3]
Поскольку никотиновые рецепторы помогают передавать исходящие сигналы для симпатической и парасимпатической систем, антагонисты никотиновых рецепторов, такие как гексаметоний, препятствуют передаче этих сигналов. Так, например, антагонисты никотиновых рецепторов мешают барорефлекс который обычно корректирует изменения артериального давления за счет симпатической и парасимпатической стимуляции сердца.[нужна цитата ]
Структура
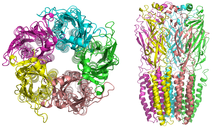
Никотиновые рецепторы с молекулярной массой 290 кДа,[9] состоят из пяти субъединиц, симметрично расположенных вокруг центрального поры.[3] Каждая субъединица включает четыре трансмембранных домена с N- и C-концом, расположенными вне клетки. Они имеют сходство с ГАМКА рецепторы, рецепторы глицина, а тип 3 рецепторы серотонина (которые все ионотропные рецепторы), или сигнатура Цис-петли белки.[10]
У позвоночных никотиновые рецепторы широко классифицируются на два подтипа в зависимости от их основных участков экспрессии: мышечный тип никотиновые рецепторы и нейронный тип никотиновые рецепторы. В рецепторах мышечного типа, обнаруженных в нервно-мышечном соединении, рецепторы представляют собой либо эмбриональную форму, состоящую из субъединиц α1, β1, γ и δ в соотношении 2: 1: 1: 1, либо взрослую форму, состоящую из α1, субъединицы β1, δ и ε в соотношении 2: 1: 1: 1.[3][4][5][11] Подтипы нейронов представляют собой различные гомомерные (все субъединицы одного типа) или гетеромерные (по крайней мере, одна α и одна β) комбинации двенадцати различных субъединиц никотиновых рецепторов: α2-α10 и β2-β4. Примеры нейронных подтипов включают: (α4)3(β2)2, (α4)2(β2)3, (α3)2(β4)3, α4α6β3 (β2)2, (α7)5, и много других. В рецепторах как мышечного, так и нейронального типа субъединицы очень похожи друг на друга, особенно в гидрофобный регионы.[нужна цитата ]
Ряд исследований с помощью электронной микроскопии и рентгеновской кристаллографии предоставили структурную информацию с очень высоким разрешением для мышечных и нейрональных nAChR и их связывающих доменов.[9][12][13][14]
Связывание с рецептором
Как и все ионные каналы, управляемые лигандами, открытие поры канала nAChR требует связывания химического посредника. Для обозначения молекул, связывающих рецепторы, используются несколько разных терминов, например: лиганд, агонист или передатчик. А также эндогенный агонист ацетилхолин, агонисты нАХР включают никотин, эпибатидин, и холин. Никотиновые антагонисты, которые блокируют рецептор, включают мекамиламин, дигидро-β-эритроидин и гексаметоний.[нужна цитата ]
В nAChR мышечного типа сайты связывания ацетилхолина расположены на границе α- или ε- или δ-субъединиц. В нейрональных nAChR сайт связывания расположен на границе α- и β-субъединицы или между двумя α-субъединицами в случае α7-рецепторов. Сайт связывания расположен во внеклеточном домене рядом с N конечная.[4][15] Когда агонист связывается с сайтом, все присутствующие субъединицы претерпевают конформационные изменения и канал открывается.[16] и открывается пора диаметром около 0,65 нм.[4]
Открытие канала
Никотиновые AChR могут существовать в различных взаимопревращаемых конформационных состояниях. Связывание агониста стабилизирует открытую и десенсибилизированный состояния. В нормальных физиологических условиях рецептору для открытия требуется ровно две молекулы ACh.[17] Открытие канала позволяет положительно заряжать ионы перемещаться по нему; особенно, натрий входит в камеру и калий выходы. Чистый поток положительно заряженных ионов направлен внутрь.
NAChR - это неселективный катионный канал, что означает, что через него могут проходить несколько различных положительно заряженных ионов.[3] Он проницаем для Na+ и K+, с некоторыми комбинациями субъединиц, которые также проницаемы для Ca2+.[4][18][19] Количество натрия и калия, которое каналы пропускают через свои поры (их проводимость ) варьируется от 50 до 110 pS, с проводимостью, зависящей от конкретного состава субъединиц, а также проникающего иона.[20]
Многие нейрональные nAChR могут влиять на высвобождение других нейротрансмиттеров.[5] Канал обычно открывается быстро и имеет тенденцию оставаться открытым до агонист распространяется прочь, что обычно занимает около 1 миллисекунда.[4] Однако AChR могут спонтанно открываться без связанных лигандов или могут спонтанно закрываться со связанными лигандами, а мутации в канале могут изменять вероятность любого события.[21][16] Следовательно, связывание ACh изменяет вероятность открытия пор, которая увеличивается по мере связывания большего количества ACh.
NAChR не может связать ACh при привязке к любому из Змеиный яд α-нейротоксины. Эти α-нейротоксины антагонистически связываются прочно и нековалентно с nAChR в скелетных мышцах и в нейронах, тем самым блокируя действие ACh на постсинаптическую мембрану, подавляя поток ионов и приводя к параличу и смерти. NAChR содержит два сайта связывания нейротоксинов змеиного яда. Прогресс в обнаружении динамики связывающего действия этих сайтов оказался трудным, хотя недавние исследования с использованием нормальный режим динамика[22] помогли предсказать природу механизмов связывания токсинов змей и ACh с nAChR. Эти исследования показали, что вращательное движение, вызванное связыванием ACh, вероятно, отвечает за открытие пор, и что одна или две молекулы α-бунгаротоксин (или другого длинноцепочечного α-нейротоксина) достаточно, чтобы остановить это движение. Токсины, кажется, связывают вместе соседние субъединицы рецептора, подавляя скручивание и, следовательно, открывающее движение.[23]
Последствия
Активация рецепторов никотином изменяет состояние нейроны через два основных механизма. С одной стороны, движение катионы вызывает деполяризация плазматической мембраны (что приводит к возбуждающий постсинаптический потенциал в нейроны ), что приводит к активации потенциалзависимые ионные каналы. С другой стороны, поступление кальция прямо или косвенно действует на различные внутриклеточные каскады. Это приводит, например, к регулированию деятельности некоторых гены или выпуск нейротрансмиттеры.[нужна цитата ]
Рецепторная регуляция
Десенсибилизация рецепторов
Связанная с лигандом десенсибилизация рецепторов была впервые охарактеризована Katz и Thesleff в никотиновых рецепторах ацетилхолина.[24]
Продолжительное или многократное воздействие стимула часто приводит к снижению чувствительности этого рецептора к стимулу, что называется десенсибилизацией. Функция nAChR может модулироваться фосфорилированием[25] путем активации зависимых от вторичных мессенджеров протеинкиназ. PKA[24] и PKC,[26] а также тирозинкиназы,[27] было показано, что он фосфорилирует nAChR, что приводит к его десенсибилизации. Сообщалось, что после продолжительного воздействия агониста на рецептор сам агонист вызывает индуцированное агонистом конформационное изменение рецептора, что приводит к десенсибилизации рецептора.[28]
Десенсибилизированные рецепторы могут вернуться в длительное открытое состояние, когда агонист связывается в присутствии положительного аллостерического модулятора, например PNU-120596.[29] Кроме того, есть данные, указывающие на то, что определенные молекулы шаперонов обладают регулирующим действием на эти рецепторы.[30]
Роли
Субъединицы никотиновых рецепторов принадлежат к мультигенному семейству (16 членов у человека), и сборка комбинаций субъединиц приводит к большому количеству различных рецепторов (для получения дополнительной информации см. База данных закрытых лигандом ионных каналов ). Эти рецепторы с очень разными кинетический, электрофизиологический и фармакологический свойства, ответить на никотин по-разному, при очень разных эффективных концентрациях. Это функциональное разнообразие позволяет им принимать участие в двух основных типах нейротрансмиссии. Классический синаптическая передача (проводная передача) включает высвобождение высоких концентраций нейромедиатора, воздействующего на непосредственно соседние рецепторы. В отличие, паракринный трансмиссия (объемная трансмиссия) предполагает нейротрансмиттеры выпущено синаптические бутоны, которые затем диффундируют через внеклеточную среду, пока не достигнут своих рецепторов, которые могут быть удалены.[31] Никотиновые рецепторы также можно найти в разных синаптических местах; например, мышечный никотиновый рецептор всегда действует постсинаптически. Нейрональные формы рецептора могут быть обнаружены как постсинаптически (участвуют в классической нейротрансмиссии), так и пресинаптически.[32] где они могут влиять на высвобождение нескольких нейротрансмиттеров.
Подразделения
Идентифицировано 17 субъединиц nAChR позвоночных, которые подразделяются на субъединицы мышечного и нейронального типа. Однако, хотя субъединица / ген α8 присутствует у видов птиц, таких как курица, она не присутствует у видов человека или млекопитающих.[33]
Субъединицы nAChR были разделены на 4 подсемейства (I-IV) на основании сходства в последовательности белков.[34] Кроме того, подсемейство III было разделено на 3 типа.
| Нейронного типа | Мышечный тип | ||||
| я | II | III | IV | ||
|---|---|---|---|---|---|
| α9, α10 | α7, α8 | 1 | 2 | 3 | α1, β1, δ, γ, ε |
| α2, α3, α4, α6 | β2, β4 | β3, α5 | |||
- α гены: CHRNA1 (мышца), CHRNA2 (нейронный), CHRNA3, CHRNA4, CHRNA5, CHRNA6, CHRNA7, CHRNA8, CHRNA9, CHRNA10
- β гены: CHRNB1 (мышца), CHRNB2 (нейронный), CHRNB3, CHRNB4
- Другие гены: CHRND (дельта), CHRNE (эпсилон), CHRNG (гамма)
Нейрональные нАХР трансмембранные белки эта форма пентамерные структуры собран из семейства субъединиц, состоящих из α2-α10 и β2-β4.[35] Эти субъединицы были обнаружены с середины 1980-х до начала 1990-х годов, когда кДНК для нескольких субъединиц nAChR были клонированы из мозга крысы и курицы, что привело к идентификации одиннадцати различных генов (двенадцать у кур), которые кодируют нейронные субъединицы nAChR; Идентифицированные гены субъединиц были названы α2 – α10 (α8 обнаружен только у кур) и β2 – β4.[36] Также было обнаружено, что различные комбинации субъединиц могут образовывать функциональные nAChR, которые могут быть активированы посредством ацетилхолин и никотин, и различные комбинации субъединиц создают подтипы nAChR с различными функциональными и фармакологическими свойствами.[37] При индивидуальной экспрессии α7, α8, α9 и α10 способны образовывать функциональные рецепторы, но для других α-субъединиц требуется присутствие β-субъединиц для образования функциональных рецепторов.[35] У млекопитающих субъединицы nAchR кодируются 17 генами, из которых девять генов, кодирующих α-субъединицы, и три кодирующие β-субъединицы, экспрессируются в головном мозге. nAChR, содержащие субъединицу β2 (β2nAChR) и α7nAChR широко экспрессируются в головном мозге, тогда как другие субъединицы nAChR имеют более ограниченную экспрессию.[38]
CHRNA5 / A3 / B4
Важный кластер генов nAchR (CHRNA5 / A3 / B4) содержит гены, кодирующие субъединицы α5, α3 и β4. Генетические исследования выявили однонуклеотидные полиморфизмы (SNP) в хромосомном локусе, кодирующем эти три гена nAChR как факторы риска никотиновая зависимость, рак легких, хроническая обструктивная болезнь легких, алкоголизм, и заболевание периферических артерий.[35][39] Гены субъединицы nAChR CHRNA5 / A3 / B4 обнаруживаются в плотном кластере в хромосомной области 15q24-25. Субъединицы nAChR, кодируемые этим локусом, образуют преобладающие подтипы никотиновых рецепторов, экспрессируемые в периферическая нервная система (ПНС) и другой ключ центральная нервная система (ЦНС) сайты, такие как медиальный хабенула, структура между лимбическим передним мозгом и средним мозгом, участвующая в основных холинергических путях.[35] Дальнейшие исследования генов CHRNA5 / A3 / B4 показали, что «нейронные» гены nAChR также экспрессируются в ненейрональных клетках, где они участвуют в различных фундаментальных процессах, таких как воспаление.[40] Гены CHRNA5 / A3 / B4 коэкспрессируются во многих типах клеток, а транскрипционная активность промоторных областей трех генов регулируется многими из одних и тех же факторов транскрипции, демонстрируя, что их кластеризация может отражать контроль экспрессии генов.[35]
CHRNA6 / CHRNB3
CHRNB3 и CHRNA6 также сгруппированы в кластер генов, расположенный на 8p11.[39] Многочисленные исследования показали, что SNPS в CHRNB3 – CHRNA6 были связаны с никотиновой зависимостью и курением, например, два SNP в CHRNB3, rs6474413 и rs10958726.[39] Генетическая изменчивость в этом регионе также влияет на восприимчивость к употреблению наркотиков, включая кокаин и алкоголь.[41] Никотиновые рецепторы, содержащие субъединицы α6 или β3, экспрессируемые в областях мозга, особенно в вентральная тегментальная область и черная субстанция, важны для употребления наркотиков из-за их роли в дофамин релиз.[42] Генетическая изменчивость этих генов может изменять чувствительность к наркотикам, вызывающим злоупотребление, множеством способов, включая изменение аминокислотной структуры белка или вызывать изменения в регуляции транскрипции и трансляции.[41]
CHRNA4 / CHRNB2
Другие хорошо изученные гены nAChR включают CHRNA4 и CHRNB2, которые были связаны как Аутосомно-доминантная ночная эпилепсия лобной доли (ADNFLE) гены.[39][43] Обе эти субъединицы nAChR присутствуют в головном мозге, и появление мутаций в этих двух субъединицах вызывает генерализованный тип эпилепсии. Примеры включают инсерционную мутацию 776ins3 CHRNA4, которая связана с ночными припадками и психическими расстройствами, и мутацию CHRNB2 I312M, которая, по-видимому, вызывает не только эпилепсию, но и очень специфические когнитивные нарушения, такие как дефицит обучения и памяти.[43][44] Между этими двумя генами существует естественная генетическая изменчивость, и анализ однонуклеотидных полиморфизмов (SNP) и других модификаций генов показывает более высокую изменчивость в гене CHRNA4, чем в гене CHRNB2, что означает, что nAChR β2, белок, кодируемый CHRNB2, ассоциируется с субъединиц больше, чем α4. CHRNA2 также упоминается как третий кандидат на ночные судороги лобных долей.[39][43]
CHRNA7
В нескольких исследованиях сообщается о связи между CHRNA7 и эндофенотипы психических расстройств и никотиновой зависимости, что способствует значительной клинической значимости α7 и проводимых исследований.[43] CHRNA7 был одним из первых генов, которые, как считалось, связаны с шизофрения. Исследования выявили несколько полиморфизмов промотора CHRNA7, которые снижают транскрипционную активность генов, что связано с шизофренией, что согласуется с обнаружением пониженных уровней nAChR a7 в мозге пациентов с шизофренией.[43] Оба подтипа nAChR, α4β2 и α7, как было обнаружено, значительно снижены в посмертных исследованиях лиц с шизофренией.[45] Кроме того, уровень курения значительно выше у больных шизофренией, что означает, что курение никотина может быть формой самолечения.[46]
Заметные вариации
Никотиновые рецепторы являются пентамерами этих субъединиц; т.е. каждый рецептор содержит пять субъединиц. Таким образом, существует огромный потенциал вариации этих субъединиц. Однако некоторые из них встречаются чаще, чем другие. Наиболее широко выраженные подтипы включают (α1)2β1δε (взрослый мышечный тип), (α3)2(β4)3 (ганглиозный тип), (α4)2(β2)3 (Типа ЦНС) и (α7)5 (другой тип ЦНС).[47] Ниже приводится сравнение:
| Рецепторного типа | Место расположения | Эффект; функции | Никотиновые агонисты | Никотиновые антагонисты |
|---|---|---|---|---|
| Мышечный тип: (α1)2β1δε[47] или же (α1)2β1δγ | Нервномышечное соединение | EPSP, в основном за счет увеличения Na+ и K+ проницаемость | ||
| Ганглиозный тип: (α3)2(β4)3 | вегетативные ганглии | EPSP, в основном за счет увеличения Na+ и K+ проницаемость | ||
| Гетеромерный тип ЦНС: (α4)2(β2)3 | Мозг | Почтовый- и пресинаптическое возбуждение,[47] в основном за счет увеличения Na+ и K+ проницаемость. Основной подтип, участвующий в повышении внимания и награждение эффекты никотина, а также патофизиология никотиновой зависимости.[49][50][51] | ||
| Далее по типу ЦНС: (α3)2(β4)3 | Мозг | Почтовый- и пресинаптическое возбуждение | ||
| Гомомерный тип ЦНС: (α7)5 | Мозг | Почтовый- и пресинаптическое возбуждение,[47] в основном за счет увеличения Na+, K+ и Ca2+ проницаемость. Основной подтип, участвующий в некоторых когнитивных эффектах никотина.[52] Более того, активация (α7)5 может улучшить нервно-сосудистый ответ при нейродегенеративном заболевании[53] и нейрогенез при ишемическом инсульте.[54] Также участвует в проангиогенных эффектах никотина и ускоряет прогрессирование хронического заболевания почек у курильщиков.[55][56][57] |
Смотрите также
- Мускариновый рецептор ацетилхолина
- Мускариновый агонист
- Мускариновый антагонист
- Никотиновый агонист
- Никотиновый антагонист
Рекомендации
- ^ Лу Б., Кван К., Левин Ю.А., Олофссон П.С., Ян Х., Ли Дж. И др. (Август 2014 г.). «Передача сигналов α7 никотинового ацетилхолинового рецептора ингибирует активацию инфламмасом, предотвращая высвобождение митохондриальной ДНК». Молекулярная медицина. 20 (1): 350–8. Дои:10.2119 / молмед.2013.00117. ЧВК 4153835. PMID 24849809.
- ^ Ямамото I (1999). «Никотин к никотиноидам: с 1962 по 1997 год». Никотиноидные инсектициды и никотиновый ацетилхолиновый рецептор. С. 3–27. Дои:10.1007/978-4-431-67933-2_1. ISBN 978-4-431-68011-6.
- ^ а б c d е ж грамм час я j k Purves D, Augustine GJ, Fitzpatrick D, Hall WC, LaMantia A, McNamara JO, White LE (2008). Неврология (4-е изд.). Sinauer Associates. стр.122 –6. ISBN 978-0-87893-697-7.
- ^ а б c d е ж Сигель Г.Дж., Агранов Б.В., Фишер С.К., Альберс Р.В., Улер М.Д. (1999). «Основы нейрохимии: молекулярные, клеточные и медицинские аспекты». Физиология и фармакология рецепторов ГАМК (6-е изд.). Американское общество нейрохимии. Получено 2008-10-01.
- ^ а б c Итиер V, Бертран Д. (август 2001 г.). «Нейрональные никотиновые рецепторы: от структуры белка к функции». Письма FEBS. 504 (3): 118–25. Дои:10.1016 / s0014-5793 (01) 02702-8. PMID 11532443.
- ^ Исии М., Курачи Ю. (1 октября 2006 г.). «Мускариновые рецепторы ацетилхолина». Текущий фармацевтический дизайн. 12 (28): 3573–81. Дои:10.2174/138161206778522056. PMID 17073660.
- ^ Лотт Э.Л., Джонс Э.Б. (2020). «Холинергическая токсичность». StatPearls. StatPearls Publishing. PMID 30969605.
- ^ Каббани Н., Нордман Дж. К., Коржиат Б. А., Велтри Д. П., Шеху А., Сеймур В. А., Адамс Д. Д. (декабрь 2013 г.). «Связаны ли никотиновые рецепторы ацетилхолина с G-белками?». BioEssays. 35 (12): 1025–34. Дои:10.1002 / bies.201300082. PMID 24185813.
- ^ а б Анвин Н. (март 2005 г.). «Уточненная структура никотинового рецептора ацетилхолина при разрешении 4А». Журнал молекулярной биологии. 346 (4): 967–89. Дои:10.1016 / j.jmb.2004.12.031. PMID 15701510.
- ^ Cascio M (май 2004 г.). «Структура и функция рецептора глицина и родственных никотиникоидных рецепторов». Журнал биологической химии. 279 (19): 19383–6. Дои:10.1074 / jbc.R300035200. PMID 15023997.
- ^ Гинятуллин Р., Нистри А., Якель Ю.Л. (июль 2005 г.). «Десенсибилизация никотиновых рецепторов ACh: формирование холинергической сигнализации». Тенденции в неврологии. 28 (7): 371–8. Дои:10.1016 / j.tins.2005.04.009. PMID 15979501.
- ^ Брейк К., ван Дейк В.Дж., Клаассен Р.В., Шуурманс М., ван дер Ост Дж., Смит А.Б., Сикма Т.К. (май 2001 г.). «Кристаллическая структура ACh-связывающего белка выявляет лиганд-связывающий домен никотиновых рецепторов». Природа. 411 (6835): 269–76. Bibcode:2001Натура.411..269Б. Дои:10.1038/35077011. PMID 11357122.
- ^ Зуридакис М., Джастас П., Заркадас Э., Хрони-Царту Д., Брегестовски П., Цартос С.Дж. (ноябрь 2014 г.). «Кристаллические структуры свободных и связанных с антагонистами состояний внеклеточного домена никотинового рецептора α9 человека». Структурная и молекулярная биология природы. 21 (11): 976–80. Дои:10.1038 / nsmb.2900. PMID 25282151.
- ^ Моралес-Перес К.Л., Новиелло К.М., Хиббс Р.Э. (октябрь 2016 г.). «Рентгеновская структура никотинового рецептора α4β2 человека». Природа. 538 (7625): 411–415. Bibcode:2016Натура.538..411М. Дои:10.1038 / природа19785. ЧВК 5161573. PMID 27698419.
- ^ Сквайр Л. (2003). Фундаментальная нейробиология (2-е изд.). Амстердам: Акад. Нажмите. п. 1426. ISBN 978-0-12-660303-3.
- ^ а б Колкухун Д., Сивилотти Л.Г. (июнь 2004 г.). «Функции и структура рецепторов глицина и некоторых их родственников». Тенденции в неврологии. 27 (6): 337–44. CiteSeerX 10.1.1.385.3809. Дои:10.1016 / j.tins.2004.04.010. PMID 15165738.
- ^ Эйдли DJ (1998). Физиология возбудимых клеток (4-е изд.). Кембридж, Великобритания: Издательство Кембриджского университета. ISBN 978-0521574150. OCLC 38067558.[страница нужна ]
- ^ Бекер Ф., Вебер М., Финк Р. Х., Адамс Д. Д. (сентябрь 2003 г.). «Активация мускариновых и никотиновых рецепторов ACh по-разному мобилизует Ca2 + в нейронах внутрисердечных ганглиев крыс». Журнал нейрофизиологии. 90 (3): 1956–64. Дои:10.1152 / ян.01079.2002. PMID 12761283.
- ^ Вебер М., Мотин Л., Галл С., Бекер Ф., Финк Р. Х., Адамс Д. Д. (январь 2005 г.). «Внутривенные анестетики ингибируют токи, опосредованные никотиновым рецептором ацетилхолина, и переходные процессы Ca2 + в нейронах внутрисердечных ганглиев крыс». Британский журнал фармакологии. 144 (1): 98–107. Дои:10.1038 / sj.bjp.0705942. ЧВК 1575970. PMID 15644873.
- ^ Мишина М., Такай Т., Имото К., Нода М., Такахаши Т., Нума С. и др. (Май 1986). «Молекулярное различие между эмбриональной и взрослой формами мышечного ацетилхолинового рецептора». Природа. 321 (6068): 406–11. Bibcode:1986Натура.321..406М. Дои:10.1038 / 321406a0. PMID 2423878.
- ^ Эйнав Т., Филлипс Р. (апрель 2017 г.). "Анализ Monod-Wyman-Changeux лиганд-зависимых мутантов ионного канала". Журнал физической химии B. 121 (15): 3813–3824. arXiv:1701.06122. Bibcode:2017arXiv170106122E. Дои:10.1021 / acs.jpcb.6b12672. ЧВК 5551692. PMID 28134524.
- ^ Левитт М., Сандер С., Стерн П.С. (февраль 1985 г.). «Нормальная динамика белков: ингибитор трипсина, крамбин, рибонуклеаза и лизоцим». Журнал молекулярной биологии. 181 (3): 423–47. Дои:10.1016 / 0022-2836 (85) 90230-х. PMID 2580101.
- ^ Самсон А.О., Левитт М. (апрель 2008 г.). «Механизм ингибирования рецептора ацетилхолина альфа-нейротоксинами, выявленный динамикой нормального режима». Биохимия. 47 (13): 4065–70. Дои:10.1021 / bi702272j. ЧВК 2750825. PMID 18327915.
- ^ а б Питчфорд С., Дэй Дж. У., Гордон А., Мочли-Розен Д. (ноябрь 1992 г.). «Десенсибилизация никотинового ацетилхолинового рецептора регулируется активацией внеклеточного аденозина». Журнал неврологии. 12 (11): 4540–4. Дои:10.1523 / JNEUROSCI.12-11-04540.1992. ЧВК 6576003. PMID 1331363.
- ^ Хуганир Р.Л., Грингард П. (февраль 1983 г.). «цАМФ-зависимая протеинкиназа фосфорилирует никотиновый ацетилхолиновый рецептор». Труды Национальной академии наук Соединенных Штатов Америки. 80 (4): 1130–4. Bibcode:1983PNAS ... 80.1130H. Дои:10.1073 / pnas.80.4.1130. ЧВК 393542. PMID 6302672.
- ^ Сафран А., Саги-Айзенберг Р., Нойман Д., Фукс С. (август 1987 г.). «Фосфорилирование ацетилхолинового рецептора протеинкиназой C и идентификация сайта фосфорилирования в дельта-субъединице рецептора». Журнал биологической химии. 262 (22): 10506–10. PMID 3038884.
- ^ Хопфилд Дж. Ф., Танк Д. В., Грингард П., Хуганир Р. Л. (декабрь 1988 г.). «Функциональная модуляция никотинового рецептора ацетилхолина путем фосфорилирования тирозина». Природа. 336 (6200): 677–80. Bibcode:1988Натура.336..677H. Дои:10.1038 / 336677a0. PMID 3200319.
- ^ Barrantes FJ (сентябрь 1978 г.). «Опосредованные агонистами изменения рецептора ацетилхолина в его мембранном окружении». Журнал молекулярной биологии. 124 (1): 1–26. Дои:10.1016/0022-2836(78)90144-4. PMID 712829.
- ^ Hurst RS, Hajós M, Raggenbass M, Wall TM, Higdon NR, Lawson JA и др. (Апрель 2005 г.). «Новый положительный аллостерический модулятор нейронального никотинового ацетилхолинового рецептора альфа7: характеристика in vitro и in vivo». Журнал неврологии. 25 (17): 4396–405. Дои:10.1523 / JNEUROSCI.5269-04.2005. ЧВК 6725110. PMID 15858066.
- ^ Садиг-Этегад С., Маджди А., Талеби М., Махмуди Дж., Бабри С. (май 2015 г.). «Регулирование никотиновых рецепторов ацетилхолина при болезни Альцгеймера: возможная роль шаперонов». Европейский журнал фармакологии. 755: 34–41. Дои:10.1016 / j.ejphar.2015.02.047. PMID 25771456.
- ^ Пиччиотто MR, Хигли MJ, Mineur YS (октябрь 2012 г.). «Ацетилхолин как нейромодулятор: холинергическая передача сигналов формирует функцию и поведение нервной системы». Нейрон. 76 (1): 116–29. Дои:10.1016 / j.neuron.2012.08.036. ЧВК 3466476. PMID 23040810.
- ^ Wonnacott S (февраль 1997 г.). «Пресинаптические никотиновые рецепторы ACh». Тенденции в неврологии. 20 (2): 92–8. Дои:10.1016 / S0166-2236 (96) 10073-4. PMID 9023878.
- ^ Грэм А., Корт Дж. А., Мартин-Руис К. М., Ярош Э., Перри Р., Фольсен С. Г. и др. (2002). «Иммуногистохимическая локализация субъединиц никотинового ацетилхолинового рецептора в мозжечке человека». Неврология. 113 (3): 493–507. Дои:10.1016 / S0306-4522 (02) 00223-3. PMID 12150770.
- ^ Le Novère N, Changeux JP (февраль 1995 г.). «Молекулярная эволюция никотинового рецептора ацетилхолина: пример мультигенного семейства в возбудимых клетках». Журнал молекулярной эволюции. 40 (2): 155–72. Bibcode:1995JMolE..40..155L. Дои:10.1007 / BF00167110. PMID 7699721.
- ^ а б c d е Improgo MR, Scofield MD, Tapper AR, Gardner PD (октябрь 2010 г.). «Генный кластер никотинового ацетилхолинового рецептора CHRNA5 / A3 / B4: двойная роль в никотиновой зависимости и раке легких». Прогресс в нейробиологии. 92 (2): 212–26. Дои:10.1016 / j.pneurobio.2010.05.003. ЧВК 2939268. PMID 20685379.
- ^ Таммимяки А., Хортон В. Дж., Стицель Дж. А. (октябрь 2011 г.). «Последние достижения в области генетической манипуляции и биологии никотиновых рецепторов ацетилхолина». Биохимическая фармакология. 82 (8): 808–19. Дои:10.1016 / j.bcp.2011.06.014. ЧВК 3162071. PMID 21704022.
- ^ Грэм А., Корт Дж. А., Мартин-Руис К. М., Ярош Э., Перри Р., Фольсен С. Г. и др. (Сентябрь 2002 г.).«Иммуногистохимическая локализация субъединиц никотинового ацетилхолинового рецептора в мозжечке человека». Неврология. 113 (3): 493–507. Дои:10.1016 / S0306-4522 (02) 00223-3. PMID 12150770.
- ^ Changeux JP (июнь 2010 г.). «Никотиновая зависимость и никотиновые рецепторы: уроки генетически модифицированных мышей». Обзоры природы. Неврология. 11 (6): 389–401. Дои:10.1038 / номер 2849. PMID 20485364.
- ^ а б c d е Гринбаум Л., Лерер Б. (октябрь 2009 г.). «Дифференциальный вклад генетической изменчивости множества никотиновых холинергических рецепторов мозга в никотиновую зависимость: недавний прогресс и новые открытые вопросы». Молекулярная психиатрия. 14 (10): 912–45. Дои:10.1038 / mp.2009.59. PMID 19564872.
- ^ Gahring LC, Rogers SW (январь 2006 г.). «Экспрессия нейронального никотинового ацетилхолинового рецептора и функция на ненейрональных клетках». Журнал AAPS. 7 (4): E885-94. Дои:10.1208 / aapsj070486. ЧВК 2750958. PMID 16594641.
- ^ а б Kamens HM, Corley RP, Richmond PA, Darlington TM, Dowell R, Hopfer CJ, et al. (Сентябрь 2016 г.). «Доказательства связи между низкочастотными вариантами CHRNA6 / CHRNB3 и антисоциальной лекарственной зависимостью». Поведенческая генетика. 46 (5): 693–704. Дои:10.1007 / s10519-016-9792-4. ЧВК 4975622. PMID 27085880.
- ^ Грэди С. Р., Салминен О., Лаверти, округ Колумбия, Уайтакер П., Макинтош Дж. М., Коллинз А.С., Маркс М.Дж. (октябрь 2007 г.). «Подтипы никотиновых рецепторов ацетилхолина на дофаминергических окончаниях полосатого тела мыши». Биохимическая фармакология. 74 (8): 1235–46. Дои:10.1016 / j.bcp.2007.07.032. ЧВК 2735219. PMID 17825262.
- ^ а б c d е Штейнлайн ОК, Бертран Д. (ноябрь 2008 г.). «Нейрональные никотиновые ацетилхолиновые рецепторы: от генетического анализа до неврологических заболеваний». Биохимическая фармакология. 76 (10): 1175–83. Дои:10.1016 / j.bcp.2008.07.012. PMID 18691557.
- ^ Бертран Д., Элмсли Ф., Хьюз Э., Траунс Дж., Сандер Т., Бертран С., Штейнлайн ОК (декабрь 2005 г.). «Мутация I312M CHRNB2 связана с эпилепсией и явными нарушениями памяти». Нейробиология болезней. 20 (3): 799–804. Дои:10.1016 / j.nbd.2005.05.013. PMID 15964197.
- ^ Бриз CR, Ли MJ, Адамс CE, Салливан Б., Логель Дж., Гиллен К.М. и др. (Октябрь 2000 г.). «Аномальная регуляция высокоаффинных никотиновых рецепторов у пациентов с шизофренией». Нейропсихофармакология. 23 (4): 351–64. Дои:10.1016 / S0893-133X (00) 00121-4. PMID 10989262.
- ^ Маклин С.Л., Грейсон Б., Идрис Н.Ф., Лесаж А.С., Пембертон Д.Д., Маки С., Нил Дж.С. (апрель 2011 г.). «Активация никотиновых рецепторов α7 улучшает вызванный фенциклидином дефицит когнитивных функций у крыс: значение для терапии когнитивной дисфункции при шизофрении». Европейская нейропсихофармакология. 21 (4): 333–43. Дои:10.1016 / j.euroneuro.2010.06.003. HDL:10454/8464. PMID 20630711.
- ^ а б c d Позвонил в HP (2003). Фармакология (5-е изд.). Эдинбург: Черчилль Ливингстон. ISBN 978-0-443-07145-4.[страница нужна ]
- ^ а б Neurosci.pharm - MBC 3320 Ацетилхолин В архиве 2007-12-27 на Wayback Machine
- ^ Сартер М (август 2015 г.). «Поведенческие и когнитивные цели для холинергического усиления». Текущее мнение в области поведенческих наук. 4: 22–26. Дои:10.1016 / j.cobeha.2015.01.004. ЧВК 5466806. PMID 28607947.
- ^ Ву Дж, Гао М., Шен Дж. Х., Ши В. X., Остер А. М., Гуткин Б. С. (октябрь 2013 г.). «Кортикальный контроль функции VTA и влияние на никотиновое вознаграждение». Биохимическая фармакология. 86 (8): 1173–80. Дои:10.1016 / j.bcp.2013.07.013. PMID 23933294.
- ^ «Никотин: биологическая активность». IUPHAR / BPS Руководство по фармакологии. Международный союз фундаментальной и клинической фармакологии. Получено 7 февраля 2016.
Kяs следующим образом; α2β4 = 9900 нМ [5], α3β2 = 14 нМ [1], α3β4 = 187 нМ [1], α4β2 = 1 нМ [4,6]. Из-за гетерогенности каналов nACh мы не пометили первичную лекарственную мишень для никотина, хотя сообщается, что α4β2 является преобладающим подтипом с высоким сродством в мозге, который опосредует никотиновую зависимость [2-3].
- ^ Левин Э.Д. (май 2012 г.). «α7-Никотиновые рецепторы и познание». Текущие цели в отношении лекарств. 13 (5): 602–6. Дои:10.2174/138945012800398937. PMID 22300026.
- ^ Садиг-Этегад С., Махмуди Дж., Бабри С., Талеби М. (ноябрь 2015 г.). «Влияние активации никотинового ацетилхолинового рецептора альфа-7 на нарушение памяти, вызванное бета-амилоидом. Возможная роль нервно-сосудистой функции». Acta Cirurgica Brasileira. 30 (11): 736–42. Дои:10.1590 / S0102-865020150110000003. PMID 26647792.
- ^ Ван Дж, Лу Зи, Фу Х, Чжан Д, Ю Л, Ли Н и др. (Май 2017). «Путь передачи сигналов никотинового рецептора альфа-7 участвует в нейрогенезе, индуцированном ChAT-положительными нейронами в субвентрикулярной зоне». Трансляционное исследование инсульта. 8 (5): 484–493. Дои:10.1007 / s12975-017-0541-7. ЧВК 5704989. PMID 28551702.
- ^ Ли Дж., Кук Дж. П. (ноябрь 2012 г.). «Никотин и патологический ангиогенез». Науки о жизни. 91 (21–22): 1058–64. Дои:10.1016 / j.lfs.2012.06.032. ЧВК 3695741. PMID 22796717.
- ^ Джайн Дж., Хаймс Э.А. (октябрь 2013 г.). «Никотиновые сигналы и прогрессирование хронической болезни почек у курильщиков». Биохимическая фармакология. 86 (8): 1215–23. Дои:10.1016 / j.bcp.2013.07.014. ЧВК 3838879. PMID 23892062.
- ^ Михалак КБ, Кэрролл Ф.И., Лютье Ч.В. (сентябрь 2006 г.). «Варениклин является частичным агонистом альфа4бета2 и полным агонистом альфа7 никотиновых рецепторов нейронов». Молекулярная фармакология. 70 (3): 801–5. Дои:10.1124 / моль.106.025130. PMID 16766716.
внешняя ссылка
 СМИ, связанные с Никотиновые рецепторы ацетилхолина в Wikimedia Commons
СМИ, связанные с Никотиновые рецепторы ацетилхолина в Wikimedia Commons- Расчетное пространственное положение никотинового ацетилхолинового рецептора в липидном бислое