Фосфатидилэтаноламин - Phosphatidylethanolamine
Фосфатидилэтаноламины являются классом фосфолипиды нашел в биологические мембраны.[1] Они синтезируются добавлением цитидин дифосфат -этаноламин к диглицериды, выпуская цитидин монофосфат. S-Аденозилметионин может впоследствии метилировать амин фосфатидилэтаноламинов с получением фосфатидилхолины. В основном его можно найти во внутреннем (цитоплазматический ) буклет липидный бислой.[2]
Функция

PtdCho - Фосфатидилхолин;
PtdEtn - Фосфатидилэтаноламин;
PtdIns -Фосфатидилинозитол;
PtdSer - Фосфатидилсерин.
В камерах
Фосфатидилэтаноламины обнаружены во всех живых клетках, составляя 25% всех фосфолипидов. В физиологии человека они обнаруживаются, в частности, в нервной ткани, такой как белое вещество из мозг, нервы, нервная ткань и спинной мозг, где они составляют 45% всех фосфолипидов.[3]
Фосфатидилэтаноламины играют роль в мембранный сплав и при разборке сократительное кольцо в течение цитокинез в деление клеток.[4] Кроме того, считается, что фосфатидилэтаноламин регулирует кривизна мембраны. Фосфатидилэтаноламин - важный прекурсор, субстрат, или донор в нескольких биологических путях.[3]
В качестве полярной головной группы фосфатидилэтаноламин создает более вязкую липидную мембрану по сравнению с фосфатидилхолин. Например, температура плавления диолеоилфосфатидилэтаноламина составляет -16 ° C, в то время как температура плавления диолеоилфосфатидилхолина составляет -20 ° C. Если бы липиды имели две пальмитоильные цепи, фосфатидилэтаноламин плавился бы при 63 ° C, тогда как фосфатидилхолин плавился бы уже при 41 ° C.[5] Более низкие температуры плавления соответствуют, в упрощенном представлении, более текучим мембранам.
В людях
Считается, что у человека метаболизм фосфатидилэтаноламина важен в сердце. Когда кровоток к сердцу ограничен, асимметричное распределение фосфатидилэтаноламина между листочками мембраны нарушается, и в результате мембрана разрушается. Кроме того, фосфатидилэтаноламин играет роль в секреции липопротеины в печени. Это потому, что пузырьки для секреции липопротеины очень низкой плотности отходит от Гольджи имеют значительно более высокую концентрацию фосфатидилэтаноламина по сравнению с другими везикулами, содержащими липопротеины очень низкой плотности.[6]Фосфатидилэтаноламин также показал способность передавать инфекционные прионы без помощи кого-либо белки или же нуклеиновые кислоты, что является его уникальной характеристикой.[7]Также считается, что фосфатидилэтаноламин играет роль в свертывании крови, поскольку он работает с фосфатидилсерин увеличить скорость тромбин формирование путем содействия привязке к Фактор V и Фактор X, два белка, которые катализируют образование тромбина из протромбин.[8] Синтез эндоканнабиноидов анандамид осуществляется из фосфатидилэтаноламина последовательным действием 2 ферментов, N-ацетилтрансфераза и фосфолипаза -D[9]
В бактериях
Где фосфатидилхолин является основным фосфолипид у животных фосфатидилэтаноламин является основным в бактерии. Одна из основных функций фосфатидилэтаноламина в бактериальных мембранах - распространение отрицательного заряда, вызванного анионный мембрана фосфолипиды. В бактерии Кишечная палочка, фосфатидилэтаноламин играет роль в поддержании пермеасы лактозы активный транспорт лактозы в клетку, а также может играть роль в других транспортных системах. Фосфатидилэтаноламин играет роль в сборке пермеазы лактозы и других мембранных белков. Он действует как «шаперон», помогая мембранным белкам правильно складывать третичные структуры чтобы они могли нормально функционировать. Когда фосфатидилэтаноламин отсутствует, транспортные белки имеют неправильную третичную структуру и не работают правильно.[10]
Фосфатидилэтаноламин также позволяет бактериальным переносчикам нескольких лекарственных препаратов функционировать должным образом и позволяет образовывать промежуточные соединения, необходимые для правильного открытия и закрытия переносчиков.[11]
Структура
Как лецитин, фосфатидилэтаноламин состоит из комбинации глицерин этерифицированный двумя жирные кислоты и фосфорная кислота. В то время как фосфатная группа сочетается с холин в фосфатидилхолине он сочетается с этаноламин в фосфатидилэтаноламине. Две жирные кислоты могут быть одинаковыми или разными и обычно находятся в положениях 1,2 (хотя они могут находиться в положениях 1,3).
Синтез
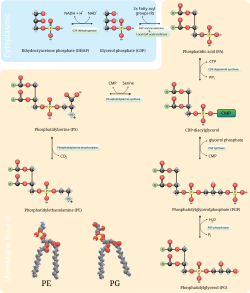
В фосфатидилсерин декарбоксилирование путь и цитидин дифосфат-этаноламин пути используются для синтеза фосфатидилэтаноламина. Фосфатидилсерин декарбоксилаза это фермент, который используется для декарбоксилирования фосфатидилсерина на первом пути. Путь декарбоксилирования фосфатидилсерина является основным источником синтеза фосфатидилэтаноламина в мембранах митохондрии. Фосфатидилэтаноламин, продуцируемый в митохондриальной мембране, также транспортируется через клетку к другим мембранам для использования. В процессе, который отражает фосфатидилхолин синтез, фосфатидилэтаноламин также производится по пути цитидиндифосфат-этаноламин с использованием этаноламин как подложка. Через несколько шагов, происходящих как в цитозоль и эндоплазматический ретикулум, путь синтеза дает конечный продукт фосфатидилэтаноламин.[12] Фосфатидилэтаноламин также в большом количестве содержится в соевом или яичном лецитине и коммерчески производится с использованием хроматографического разделения.
Регулирование
Синтез фосфатидилэтаноламина через фосфатидилсерин декарбоксилирование путь происходит быстро в внутренняя митохондриальная мембрана. Однако фосфатидилсерин производится в эндоплазматический ретикулум. Из-за этого транспорт фосфатидилсерина из эндоплазматического ретикулума к митохондриальной мембране, а затем к внутренней митохондриальной мембране ограничивает скорость синтеза по этому пути. Механизм этого транспорта в настоящее время неизвестен, но может играть роль в регуляции скорости синтеза в этом пути.[нужна цитата ]
Присутствие в пище, проблемы со здоровьем
Фосфатидилэтаноламины в пище распадаются с образованием фосфатидилэтаноламиновых соединений. Продукция Amadori как часть Реакция Майяра.[13] Эти продукты ускоряют мембрана липид перекисное окисление, вызывая окислительный стресс клеткам, которые с ними соприкасаются.[14] Окислительный стресс, как известно, вызывает ухудшение питания и ряд заболеваний. Значительные уровни продуктов Амадори-фосфатидилэтаноламина были обнаружены в самых разных пищевых продуктах, таких как шоколад, соевое молоко, детская смесь, и другие обработанные пищевые продукты. Уровни продуктов Амадори-фосфатидилэтаноламина выше в пищевых продуктах с высокими концентрациями липидов и сахара, которые имеют высокие температуры при переработке.[13] Дополнительные исследования показали, что Амадори-фосфатидилэтаноламин может играть роль в сосудистое заболевание,[15] действовать как механизм, с помощью которого сахарный диабет может увеличить заболеваемость рак,[16] и потенциально могут играть роль и в других заболеваниях. Амадори-фосфатидилэтаноламин имеет более высокую плазма концентрация у пациентов с диабетом, чем у здоровых людей, что указывает на то, что он может играть роль в развитии болезни или быть продуктом болезни.[17]
Смотрите также
Рекомендации
- ^ Веллнер, Нильс; Diep, Thi Ai; Янфельт, Кристиан; Хансен, Харальд Северин (2012). «N-ацилирование фосфатидилэтаноламина и его биологические функции у млекопитающих». Biochimica et Biophysica Acta (BBA) - Молекулярная и клеточная биология липидов. 1831 (3): 652–62. Дои:10.1016 / j.bbalip.2012.08.019. PMID 23000428.
- ^ Мишкинд, Майкл (2000). «Фосфатидилэтаноламин - в крайнем случае». Тенденции в клеточной биологии. 10 (9): 368. Дои:10.1016 / S0962-8924 (00) 01826-2.
- ^ а б Вэнс, Жан Э .; Тассева, Гергана (2012). «Образование и функции фосфатидилсерина и фосфатидилэтаноламина в клетках млекопитающих». Biochimica et Biophysica Acta (BBA) - Молекулярная и клеточная биология липидов. 1831 (3): 543–54. Дои:10.1016 / j.bbalip.2012.08.016. PMID 22960354.
- ^ Эмото, К .; Кобаяши, Т; Ямаджи, А; Aizawa, H; Яхара, я; Иноуэ, К; Умеда, М. (1996). «Перераспределение фосфатидилэтаноламина в борозде дробления делящихся клеток во время цитокинеза». Труды Национальной академии наук. 93 (23): 12867–72. Bibcode:1996PNAS ... 9312867E. Дои:10.1073 / пнас.93.23.12867. JSTOR 40713. ЧВК 24012. PMID 8917511.
- ^ См. Ссылки в Wan et al. Биохимия 47 2008[требуется проверка ]
- ^ Вэнс, Дж. Э. (2008). «Серия тематических обзоров: Глицеролипиды. Фосфатидилсерин и фосфатидилэтаноламин в клетках млекопитающих: два метаболически связанных аминофосфолипида». Журнал исследований липидов. 49 (7): 1377–87. Дои:10.1194 / мл. R700020-JLR200. PMID 18204094.
- ^ Deleault, N.R .; Piro, J. R .; Уолш, Д. Дж .; Wang, F .; Ma, J .; Geoghegan, J.C .; Супаттапон, С. (2012). «Выделение фосфатидилэтаноламина в качестве единственного кофактора для образования прионов в отсутствие нуклеиновых кислот». Труды Национальной академии наук. 109 (22): 8546–51. Bibcode:2012PNAS..109.8546D. Дои:10.1073 / pnas.1204498109. ЧВК 3365173. PMID 22586108.
- ^ Majumder, R .; Лян, X .; Quinn-Allen, M.A .; Kane, W.H .; Ленц, Б. Р. (2011). «Модуляция сборки и активности протромбиназы фосфатидилэтаноламином». Журнал биологической химии. 286 (41): 35535–42. Дои:10.1074 / jbc.M111.260141. ЧВК 3195639. PMID 21859710.
- ^ Исидро, Ф. (2014). «Каннабиноиды для лечения болезни Альцгеймера: движение к клинике». Границы фармакологии. 5: 37. Дои:10.3389 / fphar.2014.00037. ЧВК 3942876. PMID 24634659.
- ^ Кристи, W.W. (16 апреля 2012 г.). «Фосфатидилэтаноламин и родственные липиды». Липидная библиотека AOCS. Архивировано из оригинал 21 августа 2012 г.. Получено 3 сентября, 2012.
- ^ Gbaguidi, B .; Hakizimana, P .; Vandenbussche, G .; Ruysschaert, J.-M. (2007). «Конформационные изменения в бактериальном переносчике множества лекарственных средств зависят от фосфатидилэтаноламина». Клеточные и молекулярные науки о жизни. 64 (12): 1571–82. Дои:10.1007 / s00018-007-7031-0. PMID 17530171. S2CID 2078590.
- ^ Келли, Карен (28 июля 2011 г.). «Биосинтез фосфолипидов». Библиотека липидов AOCS. Получено 3 сентября, 2012.
- ^ а б Дуб, Чон-Хо; Накагава, Киётака; Миядзава, Теруо (2002). «УФ-анализ гликозилированного Амадори фосфатидилэтаноламина в пищевых продуктах и биологических образцах». Журнал исследований липидов. 43 (3): 523–9. PMID 11893788.
- ^ Дуб, Чон-Хо; Накагава, Киётака; Миядзава, Теруо (2000). «Синтетически полученный гликозилированный Амадори фосфатидилэтаноламин может запускать перекисное окисление липидов посредством свободнорадикальных реакций». Письма FEBS. 481 (1): 26–30. Дои:10.1016 / S0014-5793 (00) 01966-9. PMID 10984609. S2CID 23265125.
- ^ Дуб, Чон-Хо; Накагава, Киётака; Оикава, Шиничи; Миядзава, Теруо (2003). «Амадори-гликированный фосфатидилэтаноламин индуцирует ангиогенные дифференцировки в культивируемых эндотелиальных клетках пупочной вены человека». Письма FEBS. 555 (2): 419–23. Дои:10.1016 / S0014-5793 (03) 01237-7. PMID 14644453. S2CID 33974755.
- ^ Эйцука, Такахиро; Накагава, Киётака; Оно, Юичи; Татеваки, Наото; Нисида, Хироши; Курата, Тадао; Сёдзи, Наоки; Миядзава, Теруо (2012). «Амадори-гликированный фосфатидилэтаноламин повышает активность теломеразы в клетках карциномы поджелудочной железы человека PANC-1». Письма FEBS. 586 (16): 2542–7. Дои:10.1016 / j.febslet.2012.06.027. PMID 22750441. S2CID 5452160.
- ^ Ариидзуми, Кен; Koike, T; Охара, S; Иномата, Y; Abe, Y; Иидзима, К; Иматани, А; Ока, Т; Симосэгава, Т. (2008). «Заболеваемость рефлюкс-эзофагитом и инфекцией H.pylori у больных сахарным диабетом». Всемирный журнал гастроэнтерологии. 14 (20): 3212–7. Дои:10.3748 / wjg.14.3212. ЧВК 2712855. PMID 18506928.
внешняя ссылка
- Фосфатидилэтаноламины в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- Фосфатидилэтаноламин в липидной библиотеке AOCS.