Плазмалоген - Plasmalogen
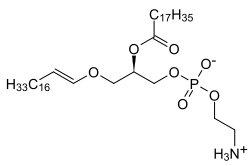
Плазмалогены являются подклассом эфирные фосфолипиды которые обычно обнаруживаются в клеточных мембранах в нервный, невосприимчивый и сердечно-сосудистая система. Есть два типа эфирные фосфолипиды, плазманил и плазменил. Плазменилфосфолипиды, которые включают плазмалоген и имеют эфирную связь в положении SN1 для алкенил группа. В отличие от плазманилфосфолипидов, они обладают эфир облигация в позиции SN1 к алкил группа.[1][2][3]
У млекопитающих SNПоложение 1 обычно происходит от жирных спиртов C16: 0, C18: 0 или C18: 1, в то время как SN2 позицию чаще всего занимают полиненасыщенные жирные кислоты (ПНЖК). Наиболее частыми головными группами, присутствующими в плазмалогенах млекопитающих, являются: этаноламин (обозначенные плазменилэталомины) или холин (обозначенные плазменилхолины).
Функции
Плазмалогены обнаружены во многих тканях человека, особенно в нервной, иммунной и сердечно-сосудистой системах.[1][2][3] В сердечной ткани человека около 30-40% холин глицерофосфолипиды плазмалогены. Еще более поразительным является тот факт, что 32% глицерофосфолипидов в сердце взрослого человека, 20% в мозге и до 70% глицерофосфолипидов этаноламина миелиновой оболочки являются плазмалогенами.[4]
Хотя функции плазмалогенов еще полностью не выяснены, было продемонстрировано, что они могут защищать клетки млекопитающих от повреждающего действия активные формы кислорода.[1][2][3] Кроме того, они были вовлечены как сигнальные молекулы и модуляторы мембранной динамики.
История
Плазмалогены были впервые описаны Фельгеном и Войтом в 1924 году на основе исследований срезов тканей.[1] Они обрабатывали эти участки ткани кислотой или хлорид ртути как часть метода окрашивания ядра. Это привело к разрыву связи плазмалоген-виниловый эфир с образованием альдегиды. В свою очередь, последний отреагировал фуксин -сернистая кислота краситель, использованный в этом методе окрашивания ядер, приводил к образованию окрашенных соединений внутри цитоплазмы клеток. Плазмалогены были названы на основании того факта, что эти окрашенные соединения присутствовали в «плазме» или внутри клетки.[1]
Биосинтез

Биосинтез плазмалогенов (ПЛ) начинается с ассоциации пероксисомальный матричные ферменты GNPAT (глицеронфосфат ацилтрансфераза) и AGPS (алкилглицеронфосфатсинтаза) на просветной стороне пероксисомальной мембраны.[5]Эти два фермента могут физически взаимодействовать друг с другом для повышения эффективности. Следовательно, фибробласты без активности AGPS имеют пониженный уровень и активность GNPAT.[6][7]
Первый этап биосинтеза катализируется GNPAT. Этот фермент ацилирует дигидроксиацетонфосфат (DHAP) в позиции sn-1. За этим следует замена ацильной группы на алкильную группу с помощью AGPS.[8]Затем 1-алкил-DHAP восстанавливается до 1-O-алкил-2-гидрокси-sn-глицерофосфата (GPA) с помощью ацил / алкил-DHAP. редуктаза расположены как в пероксисомальном, так и в эндоплазматический ретикулум (ER) мембраны.[9]Все остальные модификации происходят в ER. Есть ацил группа помещена в положение sn-2 алкил / ацил GPA ацилтрансфераза а фосфатная группа удаляется фосфатидная кислота фосфатаза с образованием 1-O-алкил-2-ацил-sn-глицерина.
Использование CDP-этаноламина a фосфотрансфераза образует 1-O-алкил-2-ацил-sn-GPEtn. После дегидрирования в положениях 1 и 2 алкильной группы с помощью системы переноса электронов и десатуразы плазманиэтаноламина окончательно образуется виниловая эфирная связь плазмалогенов. Был идентифицирован белок, соответствующий десатуразе плазманиэтаноламина и названный в бактериях CarF и PEDS1 (TMEM189). ) у людей (и животных)[10],[11].Плазменилхолин образуется из 1-O-алкил-2-ацил-sn-глицерина путем холинфосфотрансфераза. Поскольку плазмалогены холина отсутствуют, плазмалогены могут образовываться только после гидролиз из этаноламин PL в 1-O- (1Z-алкенил) -2-ацил-sn-глицерин, который может быть модифицирован холинфосфотрансферазой и холином CDP.[12][13]
Патология
Нарушения биогенеза пероксисом являются аутосомно-рецессивными заболеваниями, часто характеризующимися нарушением биосинтеза плазмалогенов. В этих случаях пероксисомальный фермент GNPAT, необходимый для начальных этапов биосинтеза плазмалогена, неправильно локализуется в цитоплазме, где он неактивен. Кроме того, генетические мутации в GNPAT или же AGPS гены могут привести к дефициту плазмалогенов, что приводит к развитию точечная ризомелическая хондродисплазия (RCDP) типа 2 или 3 соответственно.[14] В таких случаях оба экземпляра GNPAT или же AGPS ген должен быть мутирован, чтобы болезнь проявилась. В отличие от нарушений биогенеза пероксисом, другие аспекты сборки пероксисом у пациентов с RCDP2 и RCDP3 в норме, как и их способность метаболизировать жирные кислоты с очень длинной цепью. У людей с тяжелым дефицитом плазмалогенов часто наблюдаются аномалии неврологического развития, пороки развития скелета, нарушение дыхания и катаракта.[15]
Дефицит уровней плазмалогенов способствует патологии в Синдром Зеллвегера.[13]
Плазмалогеннокаутные мыши показать аналогичные изменения, такие как арест сперматогенез, развитие катаракта и дефекты центральной нервной системы миелинизация.[16][17]
Во время воспаления
Во время воспаления нейтрофилы производные миелопероксидаза производит хлорноватистая кислота (HOCl). HOCl вызывает окислительный хлорирование плазмалогенов в цепи sn-1 путем взаимодействия с виниловый эфир связь.[18] Несколько исследователей в настоящее время изучают влияние хлорированных липидов на патологию.
Возможные связи болезней
Отсутствие хороших методов для анализа плазмалогена создало трудности для ученых в оценке того, как плазмалоген может быть вовлечен в другие заболевания человека, кроме RCDP и спектра Зеллвегера, участие в которых очевидно.[13] Есть некоторые свидетельства того, что у людей низкие плазмалогены участвуют в патологии бронхолегочная дисплазия, что является важным осложнением преждевременные роды.[13] и одно исследование показывает, что уровень плазмалогенов снижается у людей с ХОБЛ кто курил по сравнению с некурящими. Есть некоторые данные, полученные от людей и животных, что в мозге снижается уровень плазмалогенов. нейродегенеративные расстройства включая болезнь Альцгеймера, болезнь Паркинсона, Болезнь Ниманна – Пика, тип C, Синдром Дауна, и рассеянный склероз, неясно, является ли это причиной или корреляцией.[13]
Эволюция
Помимо млекопитающих, плазмалогены также встречаются у беспозвоночные и одноклеточные организмы простейшие. Среди бактерии они были обнаружены у многих анаэробных видов, включая Clostridia, Megasphaera, и Veillonella. Среди аэробных бактерий плазмалогены встречаются у миксобактерий, и их десатураза плазманиэтаноламина (CarF), необходимая для образования связи простого винилового эфира, и, следовательно, плазмалогена, сохраняется как TMEM189 у людей (и животных).[19]. Было показано, что у плазмалогенов сложная эволюционная история, основанная на том факте, что их биосинтетические пути различаются у аэробных и анаэробных организмов.[20]
Недавно было продемонстрировано, что красные кровяные тельца люди и большие обезьяны (шимпанзе, бонобо, гориллы и орангутаны ) имеют различия в своем плазмалогенном составе.[3] Было обнаружено, что общий уровень плазмалогенов в эритроцитах у людей ниже, чем у бонобо, шимпанзе или горилл, но выше, чем у орангутанов. Данные по экспрессии генов всех этих видов заставили авторов предположить, что другие клетки и ткани человека и человекообразных обезьян различаются по уровням плазмалогенов. Хотя последствия этих потенциальных различий неизвестны, межвидовые различия в тканевых плазмалогенах могут влиять на функции органов и множество биологических процессов.
Рекомендации
- ^ а б c d е Nagan, N .; Зеллер, Р. А. (2001). «Плазмалогены: биосинтез и функции». Прогресс в исследованиях липидов. 40 (3): 199–229. Дои:10.1016 / S0163-7827 (01) 00003-0. PMID 11275267.
- ^ а б c Горгас, К .; Тейглер, А .; Комленович, Д .; Just, W. W. (2006). «Мышь с дефицитом эфирных липидов: отслеживание функций плазмалогена». Biochimica et Biophysica Acta (BBA) - Исследование молекулярных клеток. 1763 (12): 1511–1526. Дои:10.1016 / j.bbamcr.2006.08.038. PMID 17027098.
- ^ а б c d Moser, A. B .; Steinberg, S.J .; Уоткинс, П. А .; Moser, H.W .; Ramaswamy, K .; Siegmund, K. D .; Lee, D. R .; Ely, J. J .; Райдер, О. А .; Хасия, Дж. Г. (2011). «Эритроциты человека и обезьяны различаются по уровню и составу плазмалогенов». Липиды в здоровье и болезнях. 10: 101. Дои:10.1186 / 1476-511X-10-101. ЧВК 3129581. PMID 21679470.
- ^ Farooqui, A. A .; Хоррокс, Л. А. (2001). «Плазмалогены: рабочие липиды мембран нормальных и поврежденных нейронов и глии». Нейробиолог: обзорный журнал, посвященный нейробиологии, неврологии и психиатрии. 7 (3): 232–245. Дои:10.1177/107385840100700308. PMID 11499402. S2CID 85868497.
- ^ П. Брайтс, Х.Р. Уотерхэм, Р.Дж. Вандерс, Функции и биосинтез плазмалогенов при здоровье и болезнях, Биохим. Биофиз. Acta 1636 (2004) 219–231.
- ^ Дж. Бирманн, W.W. Просто Р.Дж. Wanders, H. Van Den Bosch, Алкил-дигидроксиацетонфосфат-синтаза и дигидроксиацетон-фосфат-ацилтрансфераза образуют белковый комплекс в пероксисомах, Eur. J. Biochem. 261 (1999) 492–499
- ^ Д. Хардеман, Х. ван ден Бош, Топография биосинтеза эфирных фосфолипидов, Biochim. Биофиз. Acta 1006 (1989) 1–8.
- ^ А.Дж. Браун, Ф. Снайдер, Алкилдигидроксиацетон-П-синтаза. Солюбилизация, частичная очистка, новый метод анализа и доказательства механизма пинг-понга, J. Biol. Chem. 257 (1982) 8835–8839
- ^ ПФ. Джеймс, А.С. Лейк, А.К. Хаджра, Л. Ларкинс, М. Робинсон, Ф. Бьюкенен, Р.А. Зеллер, Мутант клеток животных с недостаточностью активности ацил / алкилдигидроксиацетонфосфатредуктазы. Влияние на биосинтез простых эфирных и диацилглицеролипидов, J. Biol. Chem. 272 (1997) 23540–23546
- ^ Gallego-García A, Monera-Girona AJ, Pajares-Martínez E, Bastida-Martínez E, Pérez-Castaño R, Iniesta AA, Fontes M, Padmanabhan S, Elías-Arnanz M. "Реакция бактериального света выявляет орфанную десатуразу для человека. синтез плазмалогенов »(2019) Наука 366 (6461) 128-132, DOI: 10.1126 / science.aay1436, PMID = 31604315
- ^ Вернер, ER; Келлер, Массачусетс; Зайлер, S; Лакнер, К; Koch, J; Германн, М; Коассин, S; Гольдерер, G; Вернер-Фельмайер, G; Зеллер, РА; Хуло, Н; Бергер, Дж; Ватчингер, К. (7 апреля 2020 г.). "The TMEM189 ген кодирует десатуразу плазманиэтаноламина, которая вводит характерную двойную связь простого винилового эфира в плазмалогены ». Труды Национальной академии наук Соединенных Штатов Америки. 117 (14): 7792–7798. Дои:10.1073 / пнас.1917461117. PMID 32209662.
- ^ T.C. Ли, Биосинтез и возможные биологические функции плазмалогенов, Biochim. Биофиз. Acta 1394 (1998) 129–145
- ^ а б c d е Браверман, NE; Мозер, AB (сентябрь 2012 г.). «Функции плазмалогенных липидов в здоровье и болезни». Biochimica et Biophysica Acta (BBA) - Молекулярная основа болезни. 1822 (9): 1442–52. Дои:10.1016 / j.bbadis.2012.05.008. PMID 22627108.
- ^ Wanders, R .; Уотерхэм, Х. (2006). «Пероксисомальные расстройства: дефицит единственного пероксисомального фермента». Biochimica et Biophysica Acta (BBA) - Исследование молекулярных клеток. 1763 (12): 1707–20. Дои:10.1016 / j.bbamcr.2006.08.010. PMID 17055078.
- ^ Rhizomelic Chondrodysplasia Punctata Type 1. Авторы Braverman NE, Moser AB, Steinberg SJ. Редакторы в: Pagon RA, Adam MP, Bird TD, Dolan CR, Fong CT, Smith RJH, Stephens K, редакторы. Источник GeneReviews [Интернет]. Сиэтл (Вашингтон): Вашингтонский университет, Сиэтл; 1993-2014 гг. 2001 16 ноя
- ^ Горгас К., Тейглер А., Комленович Д., Just WW. Мышь с дефицитом эфирных липидов: отслеживание функций плазмалогенов. Biochim Biophys Acta. 2006 декабрь; 1763 (12): 1511-26
- ^ Родемер С., Тай Т.П., Брюггер Б., Кэрчер Т., Вернер Х., Наве К.А., Виланд Ф., Горгас К., Джаст У. Инактивация биосинтеза эфирных липидов вызывает мужское бесплодие, дефекты развития глаз и гипоплазию зрительного нерва у мышей. Hum Mol Genet. 1 августа 2003; 12 (15): 1881-95.
- ^ Альберт, Кэролайн Дж .; Кроули, Ян Р .; Сюй, Фонг-Фу; Thukkani, Arun K .; Форд, Дэвид А. (29.06.2001). «Реактивные хлорирующие виды, продуцируемые миелопероксидазой, нацелены на виниловую эфирную связь плазмалогенов. ИДЕНТИФИКАЦИЯ 2-ХЛОРОГЕКСАДЕКАНАЛА». Журнал биологической химии. 276 (26): 23733–23741. Дои:10.1074 / jbc.M101447200. ISSN 0021-9258. PMID 11301330.
- ^ Gallego-García A, Monera-Girona AJ, Pajares-Martínez E, Bastida-Martínez E, Pérez-Castaño R, Iniesta AA, Fontes M, Padmanabhan S, Elías-Arnanz M. "Реакция бактериального света выявляет орфанную десатуразу для человека. синтез плазмалогенов »(2019) Наука 366 (6461) 128-132, DOI: 10.1126 / science.aay1436, PMID = 31604315
- ^ Голдфайн, Х. (2010). «Появление, исчезновение и повторное появление плазмалогенов в эволюции». Прогресс в исследованиях липидов. 49 (4): 493–498. Дои:10.1016 / j.plipres.2010.07.003. PMID 20637230.
внешняя ссылка
- Плазмалогены в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)