Люцифераза - Luciferase
| Семейство бактериальных люциферазных монооксигеназ | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||||
| Символ | Bac_luciferase | ||||||||||
| Pfam | PF00296 | ||||||||||
| ИнтерПро | IPR016048 | ||||||||||
| PROSITE | PDOC00397 | ||||||||||
| SCOP2 | 1nfp / Объем / СУПФАМ | ||||||||||
| |||||||||||
| Каталитический домен динофлагеллят люциферазы | |||||||||
|---|---|---|---|---|---|---|---|---|---|
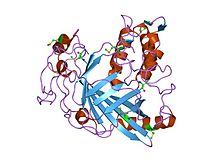 кристаллическая структура домена люциферазы из динофлагеллаты Lingulodinium polyedrum | |||||||||
| Идентификаторы | |||||||||
| Символ | Люцифераза_ кошка | ||||||||
| Pfam | PF10285 | ||||||||
| ИнтерПро | IPR018804 | ||||||||
| |||||||||
| N-концевой домен динофлагеллат люциферазы / LBP | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | Люцифераза_N | ||||||||
| Pfam | PF05295 | ||||||||
| ИнтерПро | IPR007959 | ||||||||
| |||||||||
| Домен спирального пучка динофлагеллат люциферазы | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | Люцифераза_3H | ||||||||
| Pfam | PF10284 | ||||||||
| ИнтерПро | IPR018475 | ||||||||
| |||||||||
Люцифераза это общий термин для класса окислительных ферменты которые производят биолюминесценция, и обычно отличается от фотобелок. Имя было впервые использовано Рафаэль Дюбуа кто изобрел слова люциферин и люцифераза, для субстрата и фермент, соответственно. Оба слова произошли от латинского слова Люцифер - смысл светоносец.
| Люцифераза светлячка | |||||||
|---|---|---|---|---|---|---|---|
 Структура Photinus pyralis Светлячок люцифераза. | |||||||
| Идентификаторы | |||||||
| Организм | |||||||
| Символ | Люцифераза светлячка | ||||||
| PDB | 1LCI Больше структур | ||||||
| UniProt | P08659 | ||||||
| Прочие данные | |||||||
| Номер ЕС | 1.13.12.7 | ||||||
| |||||||
Люциферазы широко используются в биотехнология, за микроскопия и, как репортерные гены, для многих из тех же приложений, что и флуоресцентные белки. Однако, в отличие от флуоресцентных белков, люциферазы не требуют внешнего источник света, но требуют добавления люциферин, расходный субстрат.
Примеры
Множество организмов регулируют свое световое производство, используя различные люциферазы в различных светоизлучающих реакциях. Большинство изученных люцифераз обнаружено у животных, в том числе светлячки, и многие морские животные, такие как копеподы, медуза, а морские анютины глазки. Однако люциферазы изучались на светящихся грибах, таких как Гриб Джек-О-Фонарь, а также примеры из других королевств, включая светящиеся бактерии, и динофлагелляты.
Светлячок и жук-щелкун
В люциферазы светлячков - из них более 2000 разновидность - и другого Elateroidea (жуки-щелкуны и родственники в целом) достаточно разнообразны, чтобы быть полезными в молекулярная филогения.[1] У светлячков необходимый кислород подается через трубку в брюшной полости, которая называется трубкой. брюшной трахея. Одна хорошо изученная люцифераза - это люцифераза Фотинини Светлячок Photinus pyralis, который имеет оптимальный pH 7,8.[2]
Морские анютины глазки
Также хорошо изучен морские анютины глазки, Renilla reniformis. В этом организме люцифераза (Ренилла-люциферин-2-монооксигеназа ) тесно связан с люциферин-связывающим белком, а также с зеленым флуоресцентным белком (GFP ). Кальций вызывает высвобождение люциферина (коэлентеразин ) из связывающего люциферин белка. Затем субстрат доступен для окисления люциферазой, где он разлагается до целентерамида с выделением энергии. В отсутствие GFP эта энергия выделялась бы в виде фотона синего света (максимальная длина волны излучения 482 нм). Однако из-за тесно связанного GFP энергия, выделяемая люциферазой, вместо этого передается через резонансная передача энергии к флуорофору GFP, а затем высвобождается в виде фотона зеленого света (максимальная длина волны излучения 510 нм). Катализируемая реакция:[3]
- коэлентеразин + О2 → целентерамид + CO2 + фотон света
Копепода
Недавно были обнаружены новые люциферазы, которые, в отличие от других люцифераз, представляют собой молекулы, секретируемые естественным путем. Одним из таких примеров является Метридия коэлентеразин -зависимая люцифераза (MetLuc, A0A1L6CBM1), который происходит от морской веслоногой рачки. Метридия длинная. В Метридия длинная секретируемый ген люциферазы кодирует белок 24 кДа, содержащий N-концевой секреторный сигнальный пептид из 17 аминокислотных остатков. Чувствительность и высокая интенсивность сигнала этой молекулы люциферазы оказались полезными во многих репортерных исследованиях. Некоторые из преимуществ использования секретируемой репортерной молекулы, такой как MetLuc, - это протокол без лизиса, который позволяет проводить анализы живых клеток и несколько анализов на одной и той же клетке.[4]
Бактериальный
Биолюминесценция бактерий наблюдается у видов Photobacterium, Вибрио фишери, Вибрио Хавейи и Вибрио Харви. Излучение света в некоторых биолюминесцентные бактерии использует «антенну», такую как «белок люмазин», чтобы принимать энергию из первичного возбужденного состояния на люциферазе, в результате чего возникает возбужденный хромофор лульназина, который излучает свет с более короткой длиной волны (более синий), в то время как в других используется желтый флуоресцентный белок (YFP) с FMN в качестве хромофора и излучает свет, смещенный в красную область по сравнению с люциферазой.[5]
Динофлагеллята
Динофлагеллята люцифераза - мульти-домен белок, состоящий из N-концевой домен, и три каталитические домены, каждому из которых предшествует домен спирального пучка. В структура динофлагеллат люциферазы каталитический домен было решено.[6] Ядро домена представляет собой 10-ниточный бета-баррель то есть структурно похожий на липокалины и FABP.[6] N-концевой домен консервированный между динофлагеллят люциферазой и люциферин связывающие белки (LBP). Было высказано предположение, что эта область может опосредовать взаимодействие между LBP и люциферазой или их ассоциацию с вакуолярный мембрана.[7]Домен спирального пучка имеет три пучок спиралей структура это содержит четыре важных гистидины которые, как считается, играют роль в pH регулирование фермент.[6] В β-стволе динофлагеллат люциферазы при pH 8 есть большой карман для размещения тетрапиррольного субстрата, но нет отверстия, позволяющего субстрату проникать. Следовательно, должно произойти значительное конформационное изменение, чтобы обеспечить доступ и пространство для лиганда в активном сайте, и источником этого изменения являются четыре N-концевых остатка гистидина.[6] При pH 8 видно, что непротонированные остатки гистидина участвуют в сети водородные связи на границе спиралей в пучке, которые блокируют доступ субстрата к активный сайт и нарушение этого взаимодействия протонирование (при pH 6,3) или заменой остатков гистидина на аланин вызывает большое молекулярное движение пучка, разделяя спирали на 11Å и открывая каталитический сайт.[6] Логично, что остатки гистидина не могут быть заменены аланином в природе, но эта экспериментальная замена дополнительно подтверждает, что более крупные остатки гистидина блокируют активный сайт. Кроме того, три последовательности Gly-Gly, одна в N-концевой спирали и две в мотиве спираль-петля-спираль, могут служить шарнирами, вокруг которых вращаются цепи, чтобы еще больше открыть путь к каталитическому сайту и увеличить активную сайт.[6]
Люцифераза динофлагеллят способна излучать свет благодаря взаимодействию со своим субстратом (люциферин ) и люциферин-связывающий белок (LBP) в сцинтилоне органелла содержится в динофлагеллятах.[6] Люцифераза действует в соответствии с люциферином и LBP, чтобы излучать свет, но каждый компонент функционирует при разном pH. Люцифераза и ее домены не активны при pH 8, но они чрезвычайно активны при оптимальном pH 6,3, тогда как LBP связывает люциферин при pH 8 и высвобождает его при pH 6,3.[6] Следовательно, люциферин высвобождается для реакции с активной люциферазой только тогда, когда сцинтиллон подкисляется до pH 6,3. Следовательно, чтобы снизить pH, закрытый по напряжению каналы в сцинтилоне мембрана открыты для входа протоны из вакуоль обладающий потенциал действия произведено механическим воздействием.[6] Следовательно, можно видеть, что потенциал действия в вакуолярной мембране приводит к подкислению, и это, в свою очередь, позволяет высвободить люциферин для взаимодействия с люциферазой в сцинтилоне, производя вспышку синего света.
Механизм реакции
Все люциферазы классифицируются как оксидоредуктазы (EC 1.13.12.- ), то есть они действуют на одиночные доноры с включением молекулярного кислорода. Поскольку люциферазы происходят из самых разных белковые семейства которые не связаны между собой, не существует объединяющего механизма, так как любой механизм зависит от комбинации люциферазы и люциферина. Однако все охарактеризованные на сегодняшний день реакции люцифераза-люциферин требуют молекулярной кислород на каком-то этапе.
Бактериальная люцифераза
Реакция, катализируемая бактериальной люциферазой, также является окислительным процессом:
- FMNH2 + O2 + RCHO → FMN + RCOOH + H2O + свет
В реакции молекулярный кислород окисляет флавинмононуклеотид и длинноцепочечный алифатический альдегид алифатическому карбоновая кислота. В результате реакции образуется возбужденный промежуточный гидроксифлавин, который дегидратируется до продукта FMN с излучением сине-зеленого света.[8]
Почти вся энергия, вкладываемая в реакцию, превращается в свет. Реакция 80%[9] до 90%[10] эффективный. Для сравнения: лампа накаливания конвертирует только около 10% своих энергия в свет[11] а светодиод 150 люмен на ватт (лм / Вт) преобразует 20% входной энергии в видимый свет.[10]
Приложения
Люциферазы можно производить в лаборатории с помощью генная инженерия для ряда целей. Люцифераза гены могут быть синтезированы и введены в организмы или трансфицированы в клетки. мышей, тутовые шелкопряды, и картофель - это всего лишь несколько организмов, которые уже были созданы для производства белка.[12]
В реакции люциферазы свет излучается, когда люцифераза воздействует на соответствующий люциферин субстрат. Эмиссия фотонов может быть обнаружена светочувствительными приборами, такими как люминометр или модифицированные оптические микроскопы. Это позволяет наблюдать за биологическими процессами.[13] Поскольку для биолюминесценции люциферазы световое возбуждение не требуется, существует минимальная автофлуоресценция и, следовательно, флуоресценция практически без фона.[14] Таким образом, даже 0,02 пг можно точно измерить с помощью стандартного сцинтилляционного счетчика.[15]
В биологических исследованиях люцифераза обычно используется в качестве репортера для оценки транскрипционный активность в клетках, трансфицированных генетической конструкцией, содержащей ген люциферазы, под контролем промоутер представляет интерес.[16] Кроме того, пролюминесцентные молекулы, которые превращаются в люциферин при активности определенного фермента, можно использовать для обнаружения активности фермента в связанных или двухэтапных анализах люциферазы. Такие подложки использовались для обнаружения каспаза деятельность и цитохром P450 деятельность, среди прочего.[13][16]
Люцифераза также может использоваться для определения уровня клеточного АТФ в жизнеспособности клеток. анализы или для анализов активности киназы.[16][17] Люцифераза может действовать как сенсорный белок АТФ через биотинилирование. Биотинилирование будет иммобилизовать люциферазу на поверхности клетки путем связывания с стрептавидин -биотин сложный. Это позволяет люциферазе обнаруживать выход АТФ из клетки и эффективно отображать высвобождение АТФ в реальном времени посредством биолюминесценции.[18] Люциферазу можно дополнительно сделать более чувствительной для обнаружения АТФ за счет увеличения интенсивности люминесценции путем изменения определенных аминокислота остатки в последовательности белка.[19]
Визуализация всего животного (обозначается как in vivo при проживании или, иначе называемом ex vivo imaging) - мощный метод изучения популяций клеток у живых животных, таких как мыши.[20] Различные типы клеток (например, стволовые клетки костного мозга, Т-клетки) могут быть сконструированы для экспрессии люциферазы, что позволяет неинвазивно визуализировать их внутри живого животного с помощью камеры чувствительного устройства с зарядовой парой (CCD камера Этот метод был использован для отслеживания туморогенеза и ответа опухолей на лечение на животных моделях.[21][22] Однако факторы окружающей среды и терапевтические вмешательства могут вызывать некоторые расхождения между опухолевой нагрузкой и интенсивностью биолюминесценции в отношении изменений пролиферативной активности. Интенсивность сигнала, измеренного с помощью визуализации in vivo, может зависеть от различных факторов, таких как абсорбция D-люциферина брюшиной, кровоток, проницаемость клеточной мембраны, наличие кофакторов, внутриклеточный pH и прозрачность вышележащих тканей в дополнение к количеству люциферазы.[23]
Люцифераза - это термочувствительный белок, который используется в исследованиях денатурация белка, проверка защитных способностей белки теплового шока. Возможности использования люциферазы продолжают расширяться.[24]
Смотрите также
Рекомендации
- ^ Гулд С.Дж., Субрамани С. (ноябрь 1988 г.). «Люцифераза светлячков как инструмент молекулярной и клеточной биологии». Аналитическая биохимия. 175 (1): 5–13. Дои:10.1016/0003-2697(88)90353-3. PMID 3072883.CS1 maint: использует параметр авторов (связь)
- ^ Стегенс Дж. П., Мин К. Л., Берненго Дж. К. (ноябрь 1998 г.). «Люцифераза светлячка имеет два нуклеотидных сайта связывания: влияние нуклеозидмонофосфата и КоА на спектры излучения света». Биохимический журнал. 336 (Pt 1) (1): 109–13. Дои:10.1042 / bj3360109. ЧВК 1219848. PMID 9806891.CS1 maint: использует параметр авторов (связь)
- ^ Шимомура О. (1985). «Биолюминесценция в море: фотобелковые системы». Симпозиумы Общества экспериментальной биологии. 39: 351–72. PMID 2871634.
- ^ Ха С., Ли Дж., Чон Э, Ким С. К., Кан Джи, Ли Дж., Ким Ю. В., Сон Ю. К., Кан Г. К., Пак Д. (июнь 2009 г.). «Клеточная система для скрининга агентов, способствующих росту волос». Архив дерматологических исследований. 301 (5): 381–85. Дои:10.1007 / s00403-009-0931-0. PMID 19277688. S2CID 23916875.
- ^ Болдуин Т.О., Кристофер Дж. А., Раушель Ф. М., Синклер Дж. Ф., Циглер М. М., Фишер А. Дж., Реймент I (декабрь 1995 г.). «Структура бактериальной люциферазы». Текущее мнение в структурной биологии. 5 (6): 798–809. Дои:10.1016 / 0959-440x (95) 80014-х. PMID 8749369.
- ^ а б c d е ж грамм час я Шульц Л.В., Лю Л., Цегельски М., Гастингс Дж. В. (февраль 2005 г.). «Кристаллическая структура люциферазы с регулируемым pH, катализирующей биолюминесцентное окисление открытого тетрапиррола». Труды Национальной академии наук Соединенных Штатов Америки. 102 (5): 1378–83. Дои:10.1073 / pnas.0409335102. ЧВК 547824. PMID 15665092.
- ^ Окамото ОК, Лю Л., Робертсон Д.Л., Гастингс Дж.В. (декабрь 2001 г.). «Члены семейства генов динофлагеллат люциферазы различаются по скорости синонимичных замен». Биохимия. 40 (51): 15862–68. CiteSeerX 10.1.1.494.3563. Дои:10.1021 / bi011651q. PMID 11747464.
- ^ Фишер А.Дж., Томпсон Т.Б., Тоден Дж.Б., Болдуин Т.О., Реймент I (сентябрь 1996 г.). «Кристаллическая структура бактериальной люциферазы с разрешением 1,5 А в условиях с низким содержанием соли». Журнал биологической химии. 271 (36): 21956–68. Дои:10.1074 / jbc.271.36.21956. PMID 8703001.
- ^ Элизабет Уилсон (18 января 1999 г.). "Что это за штука?". Новости химии и техники. 77 (3): 65. Дои:10.1021 / cen-v077n003.p065.
- ^ а б Ванесса Книветт (2009). «Освещая путь». EE Times. Архивировано из оригинал на 2012-10-05. Получено 2011-09-18.
- ^ General Electric TP-110, стр. 23, табл.
- ^ Contag CH, Bachmann MH (2002). «Достижения в визуализации биолюминесценции in vivo экспрессии генов». Ежегодный обзор биомедицинской инженерии. 4: 235–60. Дои:10.1146 / annurev.bioeng.4.111901.093336. PMID 12117758.
- ^ а б «Введение в биолюминесцентные анализы». Корпорация Промега. Архивировано из оригинал на 2010-08-14. Получено 2009-03-07.
- ^ Уильямс TM, Бурлейн Дж. Э., Огден С., Крика Л. Дж., Кант Дж. А. (январь 1989 г.). «Преимущества люциферазы светлячков как репортерного гена: приложение к промотору гена интерлейкина-2». Аналитическая биохимия. 176 (1): 28–32. Дои:10.1016/0003-2697(89)90267-4. PMID 2785354.
- ^ Нгуен В. Т., Моранж М., Бенсауд О. (июнь 1988 г.). «Люминесцентный анализ люциферазы светлячков с использованием сцинтилляционных счетчиков для количественного определения в трансфицированных клетках млекопитающих». Аналитическая биохимия. 171 (2): 404–08. Дои:10.1016/0003-2697(88)90505-2. PMID 3407940.
- ^ а б c Fan F, Wood KV (февраль 2007 г.). «Биолюминесцентные анализы для высокопроизводительного скрининга». АНАЛИЗ и технологии разработки лекарств. 5 (1): 127–36. Дои:10.1089 / adt.2006.053. PMID 17355205.
- ^ Meisenheimer PL, O'Brien MA, Cali JJ (сентябрь 2008 г.). «Субстраты люминогенных ферментов: основа новой парадигмы в дизайне анализа» (PDF). Примечания Promega. 100: 22–26. Архивировано из оригинал (PDF) на 2009-03-06. Получено 2008-10-01.
- ^ Накамура М., Миэ М., Фунабаши Х., Ямамото К., Андо Дж., Кобатаке Е. (май 2006 г.). «Обнаружение АТФ на клеточной поверхности с иммобилизованной люциферазой светлячков». Аналитическая биохимия. 352 (1): 61–67. Дои:10.1016 / j.ab.2006.02.019. PMID 16564487.
- ^ Fujii H, Noda K, Asami Y, Kuroda A, Sakata M, Tokida A (июль 2007 г.). «Повышение интенсивности биолюминесценции люциферазы светлячков с помощью генетической модификации». Аналитическая биохимия. 366 (2): 131–36. Дои:10.1016 / j.ab.2007.04.018. PMID 17540326.
- ^ Грир Л.Ф., Салай А.А. (2002). «Визуализация светового излучения от экспрессии люцифераз в живых клетках и организмах: обзор». Люминесценция. 17 (1): 43–74. Дои:10.1002 / bio.676. PMID 11816060.
- ^ Lyons SK, Meuwissen R, Krimpenfort P, Berns A (ноябрь 2003 г.). «Создание условного репортера, который обеспечивает биолюминесцентную визуализацию Cre / loxP-зависимого туморогенеза у мышей». Исследования рака. 63 (21): 7042–46. PMID 14612492.
- ^ Becher OJ, Holland EC (апрель 2006 г.). «Генно-инженерные модели имеют преимущества перед ксенотрансплантатами для доклинических исследований». Исследования рака. 66 (7): 3355–58, обсуждение 3358–59. Дои:10.1158 / 0008-5472.CAN-05-3827. PMID 16585152.
- ^ Иноуэ Й, Тодзё А., Сэкин Р., Сода Й, Кобаяши С., Номура А., Идзава К., Китамура Т., Окубо Т., Охтомо К. (май 2006 г.). «Подтверждение in vitro биолюминесцентного мониторинга прогрессирования заболевания и терапевтического ответа на модельных животных лейкемии». Европейский журнал ядерной медицины и молекулярной визуализации. 33 (5): 557–65. Дои:10.1007 / s00259-005-0048-4. PMID 16501974. S2CID 40630078.
- ^ Massoud TF, Paulmurugan R, De A, Ray P, Gambhir SS (февраль 2007 г.). «Визуализация репортерного гена белок-белковых взаимодействий у живых субъектов». Текущее мнение в области биотехнологии. 18 (1): 31–37. Дои:10.1016 / j.copbio.2007.01.007. ЧВК 4141564. PMID 17254764.
внешняя ссылка
- Обзор всей структурной информации, доступной в PDB за UniProt: P08659 (Люциферин-4-монооксигеназа) на PDBe-KB.
 СМИ, связанные с Люцифераза в Wikimedia Commons
СМИ, связанные с Люцифераза в Wikimedia Commons- Триммер Б., Заяс Р., Кази С., Льюис С., Мишель Т., Дудзински Д., Априлле Дж., Лагас С. (28 июня 2001 г.). "Светлячки и оксид азота". Университет Тафтса. Получено 2008-10-02.
- «Тенденции развития репортерных генов». reportergene.com. Получено 2009-03-07.
- "BL Web: типы люциферина". Веб-страница биолюминесценции. Калифорнийский университет в Санта-Барбаре. Получено 2009-03-07.
- «Протоколы и руководство по применению репортеров биолюминесценции». Протоколы и приложения. Корпорация Промега. Архивировано из оригинал на 08.08.2010. Получено 2009-03-07.
- «BL Web: типы люциферина». Энциклопедия науки и философии ISCID. ISCID. Архивировано из оригинал 21.09.2012. Получено 2010-04-20.
- Дэвид Гудселл. «Люцифераза». Молекула месяца. Банк данных белков. Получено 2013-01-15.