Ангиогенез - Angiogenesis
| Ангиогенез | |
|---|---|
 Ангиогенез после васкулогенеза | |
| Идентификаторы | |
| MeSH | D018919 |
| Анатомическая терминология | |

Ангиогенез это физиологический процесс, посредством которого новые кровеносный сосуд форма из уже существующих сосудов,[1][2][3] сформировался на ранней стадии васкулогенез. Ангиогенез продолжает рост сосудистая сеть процессами прорастания и расщепления.[4] Васкулогенез - это эмбриональный формирование эндотелиальный клетки из мезодерма предшественники клеток,[5] и из неоваскуляризация, хотя обсуждения не всегда точны (особенно в старых текстах). Первые сосуды в развивающихся эмбрион формируются через васкулогенез, после чего ангиогенез отвечает за большую часть, если не за весь рост кровеносных сосудов во время развитие и в болезни.[6]
Ангиогенез - это нормальный и жизненно важный процесс роста и развития, а также лечение раны и в формировании грануляционная ткань. Однако это также фундаментальный шаг в переходе от опухоли из доброкачественного состояния в злокачественный один, ведущий к использованию ингибиторы ангиогенеза в лечении рак. Существенная роль ангиогенеза в росте опухоли была впервые предложена в 1971 г. Иуда Фолкман, которые описали опухоли как «горячие и кровавые»,[7] иллюстрируя, что, по крайней мере, для многих типов опухолей, промывка перфузия и даже гиперемия характерны.
Типы
Прорастание ангиогенеза
Прорастание ангиогенеза было первой идентифицированной формой ангиогенеза, и поэтому он гораздо более понятен, чем инвагинальный ангиогенез. Это происходит в несколько хорошо известных стадий. Первоначальный сигнал исходит от участков ткани, лишенных сосудистой сети. Гипоксия, которая отмечается в этих областях, заставляет ткани требовать наличия питательных веществ и кислорода, которые позволят тканям осуществлять метаболическую активность. Из-за этого паренхиматозные клетки будут секретировать фактор роста эндотелия сосудов (VEGF-A ), который является проангиогенным фактором роста.[8] Эти биологические сигналы активируют рецепторы на эндотелиальные клетки присутствует в уже существующих кровеносных сосудах. Во-вторых, активированные эндотелиальные клетки, также известные как концевые клетки, начинают высвобождать ферменты называется протеазы которые ухудшают базальная мембрана чтобы позволить эндотелиальным клеткам вырваться из исходных (родительских) стенок сосуда. В эндотелиальные клетки тогда размножаться в окружающее матрица и образуют сплошные ростки, соединяющие соседние сосуды. Клетки, которые пролиферируют, расположены за кончиками клеток и известны как клетки стебля. Размножение этих клеток позволяет капиллярному отростку одновременно увеличиваться в длине.
По мере того, как ростки продвигаются к источнику ангиогенного стимула, эндотелиальные клетки мигрируют в тандем, используя молекулы адгезии, называемые интегрины. Эти ростки затем образуют петли, чтобы стать полноценным сосудом. просвет поскольку клетки мигрируют к месту ангиогенеза. Прорастание происходит со скоростью несколько миллиметров в день и позволяет новым сосудам расти через промежутки в сосудистая сеть. Он заметно отличается от расщепления ангиогенеза, потому что он формирует совершенно новые сосуды, а не расщепляет существующие сосуды.
Инвагинальный ангиогенез
Инвагинальный ангиогенез, также известен как расщепление ангиогенеза, представляет собой образование нового кровеносного сосуда путем разделения существующего кровеносного сосуда на два.
Инвагинация впервые наблюдалась у неонатальный крысы. В этом типе сосудистого образования стенка капилляра простирается в просвет чтобы разделить один сосуд пополам. Существует четыре фазы инвагинального ангиогенеза. Во-первых, две противоположные стенки капилляров создают зону контакта. Во-вторых, эндотелиальный клеточные соединения реорганизованы, и судно двухслойный является перфорированный позволять факторы роста и клетки для проникновения в просвет. В-третьих, между двумя новыми сосудами в зоне контакта образуется сердечник, заполненный перициты и миофибробласты. Эти клетки начинают закладывать коллаген волокна в сердцевину, чтобы обеспечить внеклеточный матрикс для роста просвета сосуда. Наконец, ядро дорабатывается без каких-либо изменений в базовой конструкции. Инвагинация важна, потому что это реорганизация существующих клеток. Это позволяет значительно увеличить количество капилляры без соответствующего увеличения количества эндотелиальные клетки. Это особенно важно при эмбриональном развитии, поскольку ресурсов недостаточно для создания богатого микрососудистая сеть с новыми клетками каждый раз, когда развивается новый сосуд.[9]
Физиология
Механическая стимуляция
Механическая стимуляция ангиогенеза изучена недостаточно. Существует значительное количество разногласий по поводу напряжение сдвига воздействуя на капилляры, вызывая ангиогенез, хотя современные знания предполагают, что усиление мышечных сокращений может усиливать ангиогенез.[10] Это может быть связано с увеличением производства оксид азота во время тренировки. Оксид азота вызывает расширение кровеносных сосудов.
Химическая стимуляция
Химическая стимуляция ангиогенеза осуществляется различными ангиогенными белками, например. интегрины и простагландины, в том числе несколько факторы роста например VEGF, FGF.
Обзор
| Стимулятор | Механизм | |
|---|---|---|
| FGF | Способствует пролиферации и дифференцировке эндотелиальных клеток, гладкомышечных клеток и фибробластов. | |
| VEGF | Влияет на проницаемость | |
| VEGFR и НРП-1 | Интегрируйте сигналы выживания | |
| Ang1 и Ang2 | Стабилизируйте сосуды | |
| PDGF (BB-гомодимер) и PDGFR | новобранец гладкомышечные клетки | |
| TGF-β, эндоглин и Рецепторы TGF-β | ↑внеклеточный матрикс производство | |
| CCL2 | Новобранцы лимфоциты на сайты воспаление | |
| Гистамин | ||
| Интегрины αVβ3, αVβ5 (?[11]) и α5β1 | Привязать матричные макромолекулы и протеиназы | |
| VE-кадгерин и CD31 | эндотелиальный соединительные молекулы | |
| эфрин | Определить образование артерий или вен | |
| активаторы плазминогена | переделывает внеклеточный матрикс, высвобождает и активирует факторы роста | |
| ингибитор активатора плазминогена-1 | стабилизирует близлежащие сосуды | |
| eNOS и СОХ-2 | ||
| AC133 | регулирует ангиобласт дифференциация | |
| ID1 /ID3 | Регулирует эндотелиальный трансдифференциация | |
| 3 класс семафорины | Модулирует адгезию, миграцию, пролиферацию и апоптоз эндотелиальных клеток. Изменяет проницаемость сосудов[12] | |
| Ного-А | Регулирует миграцию и пролиферацию эндотелиальных клеток.[13] Изменяет проницаемость сосудов.[14] |
FGF
В фактор роста фибробластов (FGF) семейство с его членами-прототипами FGF-1 (кислый FGF) и FGF-2 (основной FGF) состоит на сегодняшний день по меньшей мере из 22 известных членов.[15] Большинство из них представляют собой одноцепочечные пептиды размером 16-18 кДа и обладают высоким сродством к гепарину и гепарансульфату. В целом, FGF стимулируют множество клеточных функций путем связывания с рецепторами FGF на поверхности клетки в присутствии протеогликанов гепарина. Семейство рецепторов FGF состоит из семи членов, и все белки рецепторов являются одноцепочечными рецепторными тирозинкиназами, которые активируются посредством аутофосфорилирования, индуцированного механизмом димеризации рецепторов, опосредованной FGF. Активация рецептора вызывает каскад передачи сигнала, который приводит к активации генов и разнообразным биологическим ответам, включая дифференцировку клеток, пролиферацию и растворение матрикса, тем самым инициируя процесс митогенной активности, критической для роста эндотелиальных клеток, фибробластов и гладкомышечных клеток. .FGF-1, уникальный среди всех 22 членов семейства FGF, может связываться со всеми семью подтипами рецепторов FGF, что делает его наиболее широко действующим членом семейства FGF и мощным митогеном для различных типов клеток, необходимых для создания ангиогенного ответа в поврежденных (гипоксические) ткани, в которых происходит активация FGF-рецепторов.[16] FGF-1 стимулирует пролиферацию и дифференцировку всех типов клеток, необходимых для построения артериального сосуда, включая эндотелиальные клетки и клетки гладких мышц; этот факт отличает FGF-1 от других проангиогенных факторов роста, такие как фактор роста эндотелия сосудов (VEGF), который в первую очередь способствует образованию новых капилляров.[17][18]
Помимо FGF-1, одна из наиболее важных функций фактора роста фибробластов-2 (FGF-2 или bFGF ) способствует пролиферации эндотелиальных клеток и физической организации эндотелиальных клеток в трубчатые структуры, способствуя тем самым ангиогенезу. FGF-2 является более мощным ангиогенным фактором, чем VEGF или PDGF (фактор роста тромбоцитов ); однако он менее эффективен, чем FGF-1. Помимо стимуляции роста кровеносных сосудов, aFGF (FGF-1) и bFGF (FGF-2) играют важную роль в заживлении ран. Они стимулируют пролиферацию фибробластов и эндотелиальных клеток, которые вызывают ангиогенез и развитие грануляционной ткани; оба увеличивают кровоснабжение и заполняют раневое пространство / полость на ранних этапах процесса заживления раны.
VEGF
Фактор роста эндотелия сосудов (VEGF), как было показано, вносит основной вклад в ангиогенез, увеличивая количество капилляров в данной сети. Начальная in vitro исследования показали, что эндотелиальные клетки капилляров крупного рогатого скота будут пролиферировать и обнаруживать признаки трубчатых структур при стимуляции VEGF и bFGF, хотя результаты были более выраженными с VEGF.[19] Повышение регуляции VEGF является основным компонентом физиологической реакции на упражнения, и предполагается, что его роль в ангиогенезе является возможным лечением сосудистых повреждений.[20][21][22][23] В пробирке исследования ясно демонстрируют, что VEGF является мощным стимулятором ангиогенеза, потому что в присутствии этого фактора роста эндотелиальные клетки на пластинах будут пролиферировать и мигрировать, в конечном итоге формируя трубчатые структуры, напоминающие капилляры.[10]VEGF вызывает мощный сигнальный каскад в эндотелиальный клетки. Связывание с рецептором-2 VEGF (VEGFR-2) запускает сигнальный каскад тирозинкиназы, который стимулирует выработку факторов, которые по-разному стимулируют проницаемость сосудов (eNOS, продуцирование NO), пролиферацию / выживание (bFGF), миграцию (ICAM / VCAM / MMP) и, наконец, дифференциация в зрелые кровеносные сосуды. Механически VEGF активируется мышечными сокращениями в результате увеличения притока крови к пораженным участкам. Увеличенный поток также вызывает большое увеличение мРНК продукция рецепторов VEGF 1 и 2. Увеличение продукции рецепторов означает, что мышечные сокращения могут вызывать усиление сигнального каскада, связанного с ангиогенезом. Как часть ангиогенного сигнального каскада, NO широко рассматривается как основной участник ангиогенного ответа, поскольку ингибирование NO значительно снижает эффекты ангиогенных факторов роста. Однако ингибирование NO во время упражнений не подавляет ангиогенез, что указывает на то, что в ангиогенный ответ вовлечены и другие факторы.[10]
Ангиопоэтины
В ангиопоэтины, Ang1 и Ang2, необходимы для образования зрелых кровеносных сосудов, как показали мышиные выбить исследования.[24] Ang1 и Ang2 факторы роста белка, которые действуют путем связывания своих рецепторов, Галстук-1 и Галстук-2; в то время как это несколько спорно, кажется, что клеточные сигналы передаются в основном Галстук-2; хотя некоторые статьи показывают физиологические сигналы через Галстук-1 также. Эти рецепторы тирозинкиназы. Таким образом, они могут инициировать клеточная сигнализация когда связывание лиганда вызывает димеризацию, которая инициирует фосфорилирование на ключевые тирозины.
ММП
Еще одним важным фактором ангиогенеза является матричная металлопротеиназа (ММП). ММП помогают разрушать белки, которые поддерживают твердость стенок сосудов. Эта протеолиз позволяет эндотелиальные клетки чтобы ускользнуть в интерстициальный матрикс, как видно при прорастании ангиогенеза. Ингибирование ММП предотвращает образование новых капилляры.[25] Эти ферменты сильно регулируются в процессе формирования сосуда из-за разрушения внеклеточный матрикс снизит целостность микрососудов.[10]
DII4
Дельта-подобный лиганд 4 (DII4) представляет собой белок с отрицательным регуляторным действием на ангиогенез.[26][27] Dll4 является трансмембранным лигандом для вырезка семейства рецепторов. Было проведено много исследований, которые служили для определения последствий дельта-подобного лиганда 4. Одно исследование, в частности, оценивало влияние DII4 на васкуляризацию и рост опухоли.[28] Чтобы опухоль могла расти и развиваться, она должна иметь надлежащую сосудистую сеть. Путь VEGF жизненно важен для развития сосудистой сети, которая, в свою очередь, помогает опухолям расти. Комбинированная блокада VEGF и DII4 приводит к ингибированию прогрессирования опухоли и ангиогенеза во всей опухоли. Это происходит из-за препятствия передачи сигналов в передаче сигналов эндотелиальных клеток, которая блокирует пролиферацию и рост этих эндотелиальных клеток. Благодаря этому ингибированию клетки не растут бесконтрольно, поэтому на этом этапе рак останавливается. Однако если бы блокада была снята, клетки снова начали бы размножаться. [29]
Семафорины класса 3
Семафорины класса 3 (SEMA3s) регулируют ангиогенез путем модуляции эндотелиальная клетка адгезия, миграция, распространение, выживание и вербовка перициты.[12] Более того, семафорины могут вмешиваться в VEGF-опосредованный ангиогенез, поскольку как SEMA3, так и VEGF-A соревноваться за нейропилин связывание рецепторов в эндотелиальных клетках.[30][31] Следовательно, относительные уровни экспрессии SEMA3 и VEGF-A могут быть важны для ангиогенеза.[12]
Химическое ингибирование
An ингибитор ангиогенеза могут быть эндогенными или поступать извне как препарат, средство, медикамент или диетический компонент.
Применение в медицине
Ангиогенез как терапевтическая мишень
Ангиогенез может быть целью борьбы с такими заболеваниями, как сердечное заболевание характеризуется либо плохой васкуляризацией, либо аномальной сосудистой сетью.[32] Применение определенных соединений, которые могут ингибировать или индуцировать создание новых кровеносный сосуд в организме может помочь в борьбе с такими заболеваниями. Наличие кровеносных сосудов там, где их не должно быть, может повлиять на механические свойства ткани, увеличивая вероятность отказа. Отсутствие кровеносных сосудов в восстанавливающейся или иным образом метаболически активной ткани может препятствовать восстановлению или другим важным функциям. Некоторые заболевания, такие как ишемические хронические раны, являются результатом недостаточности или недостаточного образования кровеносных сосудов, и их можно лечить путем местного расширения кровеносных сосудов, тем самым доставляя новые питательные вещества к месту, облегчая восстановление. Другие заболевания, например, возрастные дегенерация желтого пятна, может быть вызвано локальным расширением кровеносных сосудов, нарушающим нормальные физиологические процессы.
Современное клиническое применение принципа ангиогенеза можно разделить на две основные области: антиангиогенная терапия, с которой начались ангиогенные исследования, и проангиогенная терапия. Принимая во внимание, что антиангиогенная терапия используется для борьбы с раком и злокачественными новообразованиями,[33][34] которые требуют обилия кислород и питательные вещества для пролиферации, проангиогенная терапия изучается как варианты лечения сердечно-сосудистые заболевания, причина смерти номер один в западный мир. Одним из первых применений проангиогенных методов у людей был Немецкий испытание с использованием фактора роста фибробластов 1 (FGF-1) для лечения ишемической болезни сердца.[17][35][36]
Также, что касается механизм действия проангиогенные методы можно разделить на три основные категории: генная терапия, нацеливание на интересующие гены для амплификации или ингибирования; белковая заместительная терапия, который в первую очередь управляет ангиогенными факторами роста, такими как FGF-1 или фактор роста эндотелия сосудов, VEGF; и клеточная терапия, которая включает имплантацию определенных типов клеток.
Есть еще серьезные нерешенные проблемы, связанные с генной терапией. Сложности включают эффективную интеграцию терапевтических генов в геном клеток-мишеней, снижение риска нежелательного иммунного ответа, потенциальную токсичность, иммуногенность, воспалительные реакции и онкогенез связанных с вирусными векторами, используемыми для имплантации генов, и явной сложностью генетической основы ангиогенеза. Наиболее часто встречающиеся заболевания у людей, такие как болезни сердца, высокое кровяное давление, диабет и Болезнь Альцгеймера, скорее всего, вызваны комбинированными эффектами вариаций многих генов, и, таким образом, введение одного гена может не принести значительных преимуществ при таких заболеваниях.[нужна цитата ]
Напротив, проангиогенная белковая терапия использует четко определенные, точно структурированные белки с предварительно определенными оптимальными дозами отдельного белка для болезненных состояний и с хорошо известными биологическими эффектами.[1] С другой стороны, препятствием для белковой терапии является способ доставки. Пероральный, внутривенный, внутриартериальный или внутримышечный пути введения белка не всегда столь эффективны, поскольку терапевтический белок может метаболизироваться или очищаться до того, как попадет в ткань-мишень. Клеточные проангиогенные методы лечения все еще находятся на ранней стадии исследований, и остается много открытых вопросов относительно лучших типов клеток и дозировок для использования.
Ангиогенез опухоли
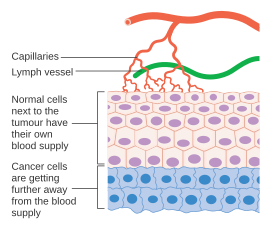
Рак клетки - это клетки, утратившие способность к контролируемому делению. А злокачественная опухоль состоит из популяции быстро делящихся и растущих раковых клеток, которые постепенно накапливаются мутации. Однако опухоли нуждаются в специальном кровоснабжении для обеспечения кислородом и другими необходимыми питательными веществами, которые им необходимы для того, чтобы вырасти до определенного размера (обычно 1-2 мм.3).[37][38]
Опухоли вызывают рост кровеносных сосудов (ангиогенез), секретируя различные факторы роста (например, VEGF ) и белки. Факторы роста, такие как bFGF и VEGF может вызвать рост капилляров в опухоли, которые, как подозревают некоторые исследователи, поставляют необходимые питательные вещества, позволяя опухоли разрастаться. В отличие от нормальных кровеносных сосудов, кровеносные сосуды опухоли расширены и имеют неправильную форму.[39] Другие клиницисты считают, что ангиогенез действительно служит отходом, убирая биологические конечные продукты, секретируемые быстро делящимися раковыми клетками. В любом случае ангиогенез является необходимым и обязательным этапом перехода от небольшого безвредного кластера клеток, который, как часто говорят, размером примерно с металлический шарик на конце шариковой ручки, к большой опухоли. Ангиогенез также необходим для распространения опухоли или метастаз. Одиночные раковые клетки могут оторваться от сформировавшейся солидной опухоли, попасть в кровеносный сосуд и быть перенесены в отдаленное место, где они могут имплантироваться и начать рост вторичной опухоли. Сейчас данные свидетельствуют о том, что кровеносный сосуд в данной солидной опухоли может быть мозаичным сосудами, состоящими из эндотелиальные клетки и опухолевые клетки. Эта мозаичность способствует значительному распространению опухолевых клеток в сосудистую сеть, что, возможно, способствует появлению циркулирующие опухолевые клетки в периферической крови больных злокачественными новообразованиями.[40] Последующий рост таких метастазов также потребует поступления питательных веществ и кислород и путь удаления отходов.
Эндотелиальные клетки долгое время считались генетически более стабильными, чем раковые клетки. Такая стабильность генома дает преимущество для нацеливания на эндотелиальные клетки с использованием антиангиогенной терапии по сравнению с химиотерапия направлено на раковые клетки, которые быстро мутируют и приобретаютустойчивость к лекарству к лечению. Именно по этой причине, эндотелиальные клетки считаются идеальной мишенью для лечения, направленного против них.[41]
Формирование кровеносных сосудов опухоли
Механизм образования кровеносных сосудов путем ангиогенеза запускается спонтанным делением опухолевых клеток в результате мутации. Затем опухолевые клетки высвобождают ангиогенные стимуляторы. Затем они перемещаются к уже установленным соседним кровеносным сосудам и активируют их рецепторы эндотелиальных клеток. Это вызывает высвобождение протеолитический ферменты сосудистой сети. Эти ферменты нацелены на определенную точку кровеносного сосуда и вызывают образование пор. Это точка, из которой будет расти новый кровеносный сосуд. Причина, по которой опухолевые клетки нуждаются в кровоснабжении, заключается в том, что они не могут расти более чем на 2–3 миллиметра в диаметре без установленного кровоснабжения, что эквивалентно примерно 50–100 клеткам.[42]
Ангиогенез при сердечно-сосудистых заболеваниях
Ангиогенез представляет собой отличную терапевтическую мишень для лечения сердечно-сосудистых заболеваний. Это мощный физиологический процесс, лежащий в основе естественной реакции нашего организма на уменьшение кровоснабжения жизненно важных органов, а именно на образование новых коллатеральных сосудов для преодоления ишемического инсульта.[17] Было проведено большое количество доклинических исследований с использованием белковой, генной и клеточной терапии на животных моделях сердечной ишемии, а также на моделях болезни периферических артерий. Воспроизводимые и достоверные успехи в этих ранних исследованиях на животных вызвали большой энтузиазм по поводу того, что этот новый терапевтический подход может быть быстро переведен на клиническую пользу для миллионов пациентов в западном мире, страдающих этими заболеваниями. Десятилетие клинических испытаний как генной, так и белковой терапии, разработанной для стимуляции ангиогенеза в недостаточно перфузируемых тканях и органах, привело, однако, от одного разочарования к другому. Хотя все эти доклинические данные, которые открывали большие перспективы для перехода терапии ангиогенеза от животных к человеку, тем или иным образом были включены в клинические испытания на ранних стадиях, FDA на сегодняшний день (2007 г.) настаивает на том, что первичные Конечной точкой для одобрения ангиогенного агента должно быть улучшение физических упражнений у пролеченных пациентов.[43]
Эти неудачи предполагали, что либо это неправильные молекулярные мишени для индукции неоваскуляризации, либо что их можно эффективно использовать только при правильном составлении и применении, либо что их презентация в контексте общей клеточной микросреды могут играть жизненно важную роль в их полезности. Может возникнуть необходимость представить эти белки таким образом, чтобы имитировать естественные сигнальные события, включая концентрация, пространственный и временный профили и их одновременное или последовательное представление с другими соответствующими факторами.[44]
Упражнение
Ангиогенез обычно связан с упражнение аэробики и упражнения на выносливость. В то время как артериогенез производит сетевые изменения, которые позволяют значительно увеличить объем общего потока в сети, ангиогенез вызывает изменения, которые обеспечивают большую доставку питательных веществ в течение длительного периода времени. Капилляры предназначены для обеспечения максимальной эффективности доставки питательных веществ, поэтому увеличение количества капилляров позволяет сети доставлять больше питательных веществ за то же время. Большее количество капилляров также способствует большему обмену кислорода в сети. Это жизненно важно для тренировок на выносливость, поскольку позволяет человеку продолжать тренировки в течение длительного периода времени. Однако нет экспериментальных данных, свидетельствующих о том, что при упражнениях на выносливость требуется повышенная капиллярность для увеличения максимальной доставки кислорода.[10]
Дегенерация желтого пятна
Сверхэкспрессия VEGF вызывает повышенную проницаемость кровеносных сосудов в дополнение к стимуляции ангиогенеза. Во влажном дегенерация желтого пятна, VEGF вызывает пролиферацию капилляров в сетчатку. Поскольку усиление ангиогенеза также вызывает отек кровь и другие жидкости сетчатки попадают в сетчатка, вызывая потерю зрения. Антиангиогенные препараты, нацеленные на пути VEGF, теперь успешно используются для лечения этого типа дегенерации желтого пятна.
Тканевые инженерные конструкции
Важен ангиогенез сосудов от тела хозяина к имплантированным тканевым конструкциям. Успешная интеграция часто зависит от тщательной васкуляризации конструкции, поскольку она обеспечивает кислород и питательные вещества и предотвращает некроз в центральных областях имплантата.[45] Было показано, что PDGF стабилизирует васкуляризацию коллаген-гликозаминогликановых каркасов.[46]
Количественная оценка
Количественная оценка параметров сосудистой сети, таких как плотность микрососудов, сопряжена с различными осложнениями из-за предпочтительного окрашивания или ограниченного представления тканей на гистологических срезах. Недавние исследования показали полную трехмерную реконструкцию сосудистой структуры опухоли и количественную оценку сосудистых структур целых опухолей на животных моделях.[47]
Смотрите также
использованная литература
- ^ а б Понимание ангиогенеза из систематического обзора. Нью-Йорк: Nova Science. 2013. ISBN 978-1-62618-114-4.
- ^ Бирбрайр А., Чжан Т., Ван З.М., Месси М.Л., Минц А., Дельбоно О. (январь 2015 г.). «Перициты на стыке регенерации тканей и патологии». Клиническая наука. 128 (2): 81–93. Дои:10.1042 / CS20140278. ЧВК 4200531. PMID 25236972.
- ^ Бирбрайр А., Чжан Т., Ван З.М., Месси М.Л., Олсон Д.Д., Минц А., Дельбоно О. (июль 2014 г.). «Перициты 2 типа участвуют в нормальном и опухолевом ангиогенезе». Американский журнал физиологии. Клеточная физиология. 307 (1): C25-38. Дои:10.1152 / ajpcell.00084.2014. ЧВК 4080181. PMID 24788248.
- ^ «Развитие лимфатической сосудистой сети» (PDF). www.columbia.edu. Получено 17 декабря 2018.
- ^ Рисау В., Пламя I (1995). «Васкулогенез». Ежегодный обзор клеточной биологии и биологии развития. 11: 73–91. Дои:10.1146 / annurev.cb.11.110195.000445. PMID 8689573.
- ^ Flamme I, Frölich T., Risau W (ноябрь 1997 г.). «Молекулярные механизмы васкулогенеза и эмбрионального ангиогенеза». Журнал клеточной физиологии. 173 (2): 206–10. Дои:10.1002 / (SICI) 1097-4652 (199711) 173: 2 <206 :: AID-JCP22> 3.0.CO; 2-C. PMID 9365523.
- ^ Джон С. Пенн (11 марта 2008 г.). Сетчатка и хориоидальный ангиогенез. Springer. С. 119–. ISBN 978-1-4020-6779-2. Получено 26 июн 2010.
- ^ Adair TH, Montani JP. Ангиогенез. Сан-Рафаэль (Калифорния): Morgan & Claypool Life Sciences; 2010. Глава 1, Обзор ангиогенеза. Доступна с: https://www.ncbi.nlm.nih.gov/books/NBK53238/
- ^ Бурри PH, Глущук Р., Джонов В. (ноябрь 2004 г.). «Инвагинальный ангиогенез: его возникновение, характеристики и значение». Динамика развития. 231 (3): 474–88. Дои:10.1002 / dvdy.20184. PMID 15376313. S2CID 35018922.
- ^ а б c d е Приор Б.М., Ян Х.Т., Терджунг Р.Л. (сентябрь 2004 г.). «Что заставляет сосуды расти при тренировках?». Журнал прикладной физиологии. 97 (3): 1119–28. Дои:10.1152 / japplphysiol.00035.2004. PMID 15333630.
- ^ Возможно ингибитор ангиогенеза: Шеппард Д. (октябрь 2002 г.). «Эндотелиальные интегрины и ангиогенез: уже не так просто». Журнал клинических исследований. 110 (7): 913–4. Дои:10.1172 / JCI16713. ЧВК 151161. PMID 12370267.
- ^ а б c Mecollari V, Nieuwenhuis B, Verhaagen J (2014). «Перспектива роли передачи сигналов семафорина класса III в травме центральной нервной системы». Границы клеточной неврологии. 8: 328. Дои:10.3389 / fncel.2014.00328. ЧВК 4209881. PMID 25386118.
- ^ Руст, Руслан; Грённерт, Лиза; Гантнер, Кристина; Энцлер, Алинда; Малдерс, Гертье; Вебер, Ребекка З .; Сиверт, Артур; Лимасале, Януар Д. П .; Мейнхардт, Андреа; Maurer, Michael A .; Сартори, Андреа М .; Хофер, Анна-Софи; Вернер, Карстен; Шваб, Мартин Э. (9 июля 2019 г.). «Таргетная терапия Nogo-A способствует восстановлению сосудов и функциональному восстановлению после инсульта». Труды Национальной академии наук. 116 (28): 14270–14279. Дои:10.1073 / пнас.1905309116. ЧВК 6628809. PMID 31235580.
- ^ Руст, Руслан; Вебер, Ребекка З .; Грённерт, Лиза; Малдерс, Гертье; Maurer, Michael A .; Хофер, Анна-Софи; Сартори, Андреа М .; Шваб, Мартин Э. (27 декабря 2019 г.). «Антитела против Nogo-A предотвращают протекание сосудов и действуют как проангиогенные факторы после инсульта». Научные отчеты. 9 (1): 20040. Bibcode:2019НатСР ... 920040Р. Дои:10.1038 / s41598-019-56634-1. ЧВК 6934709. PMID 31882970.
- ^ Орниц Д.М., Ито Н. (2001). «Факторы роста фибробластов». Геномная биология. 2 (3): ОБЗОРЫ 3005. Дои:10.1186 / gb-2001-2-3-reviews3005. ЧВК 138918. PMID 11276432.
- ^ Blaber M, DiSalvo J, Thomas KA (февраль 1996 г.). «Рентгеновская кристаллическая структура фактора роста кислых фибробластов человека». Биохимия. 35 (7): 2086–94. CiteSeerX 10.1.1.660.7607. Дои:10.1021 / bi9521755. PMID 8652550.
- ^ а б c Стегманн Т.Дж. (декабрь 1998 г.). «FGF-1: фактор роста человека в индукции неоангиогенеза». Заключение эксперта по исследуемым препаратам. 7 (12): 2011–5. Дои:10.1517/13543784.7.12.2011. PMID 15991943.
- ^ Хурана Р., Саймонс М. (апрель 2003 г.). «Выводы из испытаний ангиогенеза с использованием фактора роста фибробластов для прогрессирующего артериосклеротического заболевания». Тенденции в сердечно-сосудистой медицине. 13 (3): 116–22. Дои:10.1016 / S1050-1738 (02) 00259-1. PMID 12691676.
- ^ Гото Ф., Гото К., Вайндель К., Фолкман Дж. (Ноябрь 1993 г.). «Синергетические эффекты фактора роста эндотелия сосудов и основного фактора роста фибробластов на пролиферацию и образование пуповины эндотелиальных клеток бычьих капилляров в коллагеновых гелях». Лабораторные исследования; Журнал технических методов и патологии. 69 (5): 508–17. PMID 8246443.
- ^ Ding YH, Luan XD, Li J, Rafols JA, Guthinkonda M, Diaz FG, Ding Y (декабрь 2004 г.). «Избыточная экспрессия ангиогенных факторов, вызванная физическими упражнениями, и снижение ишемии / реперфузионного повреждения при инсульте». Текущие нейрососудистые исследования. 1 (5): 411–20. Дои:10.2174/1567202043361875. PMID 16181089. Архивировано из оригинал 19 апреля 2012 г.
- ^ Гэвин Т.П., Робинсон С.Б., Йегер Р.К., Англия Дж. А., Нифонг Л. В., Хикнер Р. К. (январь 2004 г.). «Ответ ангиогенного фактора роста на острые системные упражнения в скелетных мышцах человека». Журнал прикладной физиологии. 96 (1): 19–24. Дои:10.1152 / japplphysiol.00748.2003. PMID 12949011. S2CID 12750224.
- ^ Краус Р.М., Столлингс Х.В., Йегер Р.К., Гэвин Т.П. (апрель 2004 г.). «Циркулирующая плазменная реакция VEGF на упражнения у мужчин, ведущих малоподвижный образ жизни и тренированных на выносливость». Журнал прикладной физиологии. 96 (4): 1445–50. Дои:10.1152 / japplphysiol.01031.2003. PMID 14660505.
- ^ Ллойд П.Г., Приор Б.М., Ян Х.Т., Терджунг Р.Л. (май 2003 г.). «Экспрессия ангиогенного фактора роста в скелетных мышцах крысы в ответ на тренировку». Американский журнал физиологии. Сердце и физиология кровообращения. 284 (5): H1668-78. Дои:10.1152 / ajpheart.00743.2002. PMID 12543634.
- ^ Терстон G (октябрь 2003 г.). «Роль ангиопоэтинов и тирозинкиназ рецепторов Tie в ангиогенезе и лимфангиогенезе». Исследования клеток и тканей. 314 (1): 61–8. Дои:10.1007 / s00441-003-0749-6. PMID 12915980. S2CID 2529783.
- ^ Хаас Т.Л., Милкевич М., Дэвис С.Дж., Чжоу А.Л., Эггинтон С., Браун М.Д., Мадри Дж.А., Худличка О. (октябрь 2000 г.). «Активность матричной металлопротеиназы необходима для индуцированного активностью ангиогенеза в скелетных мышцах крысы». Американский журнал физиологии. Сердце и физиология кровообращения. 279 (4): H1540-7. Дои:10.1152 / ajpheart.2000.279.4.H1540. PMID 11009439. S2CID 2543076.
- ^ Лобов И.Б., Ренар Р.А., Пападопулос Н., Гейл Н.В., Терстон Г., Янкопулос Г.Д., Виганд С.Дж. (февраль 2007 г.). «Дельта-подобный лиганд 4 (Dll4) индуцируется VEGF как негативный регулятор ангиогенного разрастания». Труды Национальной академии наук Соединенных Штатов Америки. 104 (9): 3219–24. Bibcode:2007PNAS..104.3219L. Дои:10.1073 / pnas.0611206104. ЧВК 1805530. PMID 17296940.
- ^ Hellström M, Phng LK, Hofmann JJ, Wallgard E, Coultas L, Lindblom P, Alva J, Nilsson AK, Karlsson L, Gaiano N, Yoon K, Rossant J, Iruela-Arispe ML, Kalén M, Gerhardt H, Betsholtz C ( Февраль 2007 г.). «Передача сигналов Dll4 через Notch1 регулирует образование концевых клеток во время ангиогенеза». Природа. 445 (7129): 776–80. Bibcode:2007Натура.445..776H. Дои:10.1038 / природа05571. PMID 17259973. S2CID 4407198.
- ^ Сегарра М., Уильямс К.К., Сьерра М.Л., Бернарндо М., Маккормик П.Дж., Мерик Д., Регино С., Чойк П., Тосато Г. 2008. Активация Dll4 передачи сигналов Notch снижает васкуляризацию опухоли и ингибирует рост опухоли. Журнал крови. 112 (5): 1904-1911.
- ^ Ли Ди, Ким Ди, Бин Чой Й, Кан К., Сунг Э.С., Ан ДжХ, Гу Дж, Йом Д.Х., Сук Чан Х, Дук Мун К., Хун Ли С., Ю В.К. (2016). «Одновременная блокада VEGF и Dll4 HD105, биспецифическим антителом, ингибирует прогрессирование опухоли и ангиогенез». mAbs. 8 (5): 892–904. Дои:10.1080/19420862.2016.1171432. ЧВК 4968104. PMID 27049350.
- ^ Сокер С., Такашима С., Miao HQ, Neufeld G, Klagsbrun M (март 1998 г.). «Нейропилин-1 экспрессируется эндотелиальными и опухолевыми клетками в качестве изоформ-специфичного рецептора фактора роста эндотелия сосудов». Ячейка. 92 (6): 735–45. Дои:10.1016 / s0092-8674 (00) 81402-6. PMID 9529250. S2CID 547080.
- ^ Херцог Б., Pellet-Many C, Бриттон Г., Харцулакис Б., Захари И.К. (август 2011 г.). «Связывание VEGF с NRP1 необходимо для стимуляции VEGF миграции эндотелиальных клеток, образования комплекса между NRP1 и VEGFR2 и передачи сигналов через фосфорилирование FAK Tyr407». Молекулярная биология клетки. 22 (15): 2766–76. Дои:10.1091 / mbc.E09-12-1061. ЧВК 3145551. PMID 21653826.
- ^ Феррара Н., Кербель Р.С. (декабрь 2005 г.). «Ангиогенез как терапевтическая мишень». Природа. 438 (7070): 967–74. Bibcode:2005Натура.438..967F. Дои:10.1038 / природа04483. PMID 16355214. S2CID 1183610.
- ^ Folkman J, Klagsbrun M (январь 1987 г.). «Ангиогенные факторы». Наука. 235 (4787): 442–7. Bibcode:1987Наука ... 235..442F. Дои:10.1126 / science.2432664. PMID 2432664.
- ^ Фолкман Дж (сентябрь 1996 г.). «Борьба с раком путем атаки на его кровоснабжение». Scientific American. 275 (3): 150–4. Bibcode:1996SciAm.275c.150F. Дои:10.1038 / scientificamerican0996-150. PMID 8701285.
- ^ Стегманн Т.Дж., Хопперт Т., Шнайдер А., Гемейнхардт С., Кохер М., Ибинг Р., Штрупп Г. (сентябрь 2000 г.). «[Индукция миокардиального неоангиогенеза факторами роста человека. Новый терапевтический подход к ишемической болезни сердца]». Herz (на немецком). 25 (6): 589–99. Дои:10.1007 / PL00001972. PMID 11076317.
- ^ Фолкман Дж (февраль 1998 г.). «Ангиогенная терапия сердца человека». Тираж. 97 (7): 628–9. Дои:10.1161 / 01.CIR.97.7.628. PMID 9495294.
- ^ Макдугалл С.Р., Андерсон А.Р., капеллан Массачусетса (август 2006 г.). «Математическое моделирование динамического адаптивного ангиогенеза, индуцированного опухолью: клиническое значение и стратегии терапевтического воздействия». Журнал теоретической биологии. 241 (3): 564–89. Дои:10.1016 / j.jtbi.2005.12.022. PMID 16487543.
- ^ Разлив F, Герреро П., Аларкон Т., Майни П.К., Бирн Х.М. (февраль 2015 г.). «Мезоскопическое и континуальное моделирование ангиогенеза». Журнал математической биологии. 70 (3): 485–532. arXiv:1401.5701. Дои:10.1007 / s00285-014-0771-1. ЧВК 5320864. PMID 24615007.
- ^ Гонсалес-Перес Р. Р., Руэда Б. Р. (2013). Регуляторы ангиогенеза опухоли (первое изд.). Бока-Ратон: Тейлор и Фрэнсис. п. 347. ISBN 978-1-4665-8097-8. Получено 2 октября 2014.
- ^ Аллард В.Дж., Матера Дж., Миллер М.К., Реполлет М., Коннелли М.С., Рао С., Тиббе А.Г., Ур Дж. В., Терстаппен Л. В. (октябрь 2004 г.). «Опухолевые клетки циркулируют в периферической крови всех основных карцином, но не у здоровых субъектов или пациентов с незлокачественными заболеваниями». Клинические исследования рака. 10 (20): 6897–904. Дои:10.1158 / 1078-0432.CCR-04-0378. PMID 15501967.
- ^ Багри А., Курос-Мехр Х., Леонг К.Г., Пахарь Г.Д. (март 2010 г.). «Использование адъювантной терапии против VEGF при раке: проблемы и обоснование». Тенденции в молекулярной медицине. 16 (3): 122–32. Дои:10.1016 / j.molmed.2010.01.004. PMID 20189876.
- ^ Нисида Н., Яно Х., Нишида Т., Камура Т., Кодзиро М. (сентябрь 2006 г.). «Ангиогенез при раке». Здоровье сосудов и управление рисками. 2 (3): 213–9. Дои:10.2147 / vhrm.2006.2.3.213. ЧВК 1993983. PMID 17326328.
- ^ Hariawala MD, Sellke FW (июнь 1997 г.). «Ангиогенез и сердце: терапевтическое значение». Журнал Королевского медицинского общества. 90 (6): 307–11. Дои:10.1177/014107689709000604. ЧВК 1296305. PMID 9227376.
- ^ Cao L, Mooney DJ (ноябрь 2007 г.). «Пространственно-временной контроль передачи сигналов фактора роста для терапевтической неоваскуляризации». Расширенные обзоры доставки лекарств. 59 (13): 1340–50. Дои:10.1016 / j.addr.2007.08.012. ЧВК 2581871. PMID 17868951.
- ^ Роукема, Йерун; Хадемхоссейни, Али (сентябрь 2016 г.). «Васкуляризация и ангиогенез в тканевой инженерии: помимо создания статических сетей». Тенденции в биотехнологии. 34 (9): 733–745. Дои:10.1016 / j.tibtech.2016.03.002. PMID 27032730.
- ^ Амарал, Роналду Хосе Фариас Корреа; Кавана, Брентон; О'Брайен, Фергал Джозеф; Кирни, Катал Джон (16 декабря 2018 г.). «Фактор роста, полученный из тромбоцитов, стабилизирует васкуляризацию коллаген-гликозаминогликановых каркасов». Журнал тканевой инженерии и регенеративной медицины. 13 (2): 261–273. Дои:10.1002 / термин.2789. PMID 30554484. S2CID 58767660.
- ^ Чиа-Чи Чиен; Иван М. Кемпсон; Ченг Лян Ван; Х. Х. Чен; Юкуан Хуа; N. Y. Chen; Т. К. Ли; Кельвин К.-К. Цай; Минг-Шэн Лю; Kwang-Yu Change; К. С. Ян; Г. Маргаритондо (май – июнь 2013 г.). «Полное микромасштабное профилирование микроангиогенеза опухоли». Достижения биотехнологии. 31 (3): 396–401. Дои:10.1016 / j.biotechadv.2011.12.001. PMID 22193280.
внешние ссылки
- Ангиогенез при сердечных заболеваниях от ангиопластики.Org
- Ангиогенез - Виртуальная библиотека биохимии, молекулярной биологии и клеточной биологии
- Визуализация ангиогенеза с помощью GFP
- NCI Понимание рака серии по ангиогенезу
- Adair, TH; Монтани, JP (2010). Ангиогенез. Сан-Рафаэль: Науки о жизни Моргана и Клейпула. PMID 21452444. Учебник по теме находится в свободном доступе в NCBI.