Аденилосукцинатлиаза - Adenylosuccinate lyase




| Аденилосукцинатлиаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
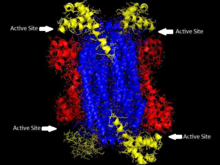 'Гомотетрамерная структура ASL в Thermotoga maritima Домен 1 красный, Домен 2 выделен синим цветом, Домен 3 выделен желтым. Эта структура была вдохновлена статьей Тота и Йейтса.[5] | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 4.3.2.2 | ||||||||
| Количество CAS | 9027-81-0 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
Аденилосукцинатлиаза (или же аденилосукциназа) является фермент что у людей кодируется ADSL ген.[6]
Аденилосукцинатлиаза превращает аденилосукцинат к AMP и фумарат как часть пуриновый нуклеотидный цикл. ASL катализирует две реакции в пути биосинтеза пурина, который производит AMP; ASL расщепляет аденилосукцинат до AMP и фумарат, и раскалывает SAICAR в AICAR и фумарат.
Аденилосукцинатлиаза входит в состав β-устранение суперсемейство ферментов и проходит через E1cb механизм реакции. Фермент - это гомотетрамер по три домена в каждом мономер и четыре активные сайты на гомотетрамер.
Точечные мутации в аденилосукцинате, что снижает ферментативная активность вызвать клинические симптомы, которые отмечают состояние дефицит аденилосукцинатлиазы.
Этот белок может использовать морфеин модель аллостерическая регуляция.[7]
Функция

Аденилосукцинатлиаза (ASL) - это фермент, который катализирует две реакции в de novo биосинтетический пурин путь. В обеих реакциях он использует Реакция элиминирования E1cb механизм отщепления фумарата от субстрата. В первой реакции ASL превращает 5-аминоимидазол- (N-сукцинилокарбоксамид) риботид (SAICAR) в 5-аминоимидазол-4-карбоксамид риботид (AICAR) и фумарат. AICAR проходит еще три реакции, прежде чем превратится в аденилосукцинат (также называемый сукциниладенозинмонофосфатом или SAMP), который затем расщепляется на аденозинмонофосфат (AMP) и фумарат.[8] ASL важен для клеток не только из-за его участия в создании пуринов, необходимых для клеточная репликация, но также потому, что он помогает регулировать метаболические процессы, контролируя уровни AMP и фумарата в клетке.[9]

Структура
Подразделения
Аденилосукцинатлиаза принадлежит к суперсемейству β-элиминирования и по своей структуре представляет собой гомотетрамер. Мономер аденилосукцинатлиазы имеет три домена. В Thermotoga maritima, домен 1 содержит 7 α-спирали в остатки 1-93, включая His68, который является высококонсервативным и ранее считался каталитическим кислота в активный сайт.[5] Более поздние исследования показали, что His171 в домене 2, который ранее считался каталитическим основание, может фактически действовать как каталитическая кислота, по крайней мере, в кишечная палочка.[9] Домен 2 состоит из остатков 94-341 и содержит 5 α-спиралей и только мономер. β-лист. Домен 3 состоит из 7 α-спиралей. Ядро тетрамера состоит из четырех копий домена 2, и есть две копии каждого из доменов 1 и 3 на каждом конце тетрамера, дающие тетрамер D2 двугранная симметрия. Тетрамер имеет четыре активных сайта, на каждом из которых встречаются три домена.[5]
Аденилосукцинатлиаза у людей и Bacillus subtilis возможно конкурентно подавленный посредством аналог субстрата аденозинфосфонамасляная кислота 2 ’(3’), 5’-дифосфат (APBADP). APBADP является конкурентным ингибитором обеих реакций, катализируемых аденилосукцинатлиазой, и кинетические исследования с APBADP показывают, что субстраты для обеих реакций используют один и тот же активный центр.[10] В реакции, катализируемой ASL, расщепляющей аденилосукцинат на аденозинмонофосфат (AMP) и фумарат, AMP должен немного вращаться после завершения реакции и до высвобождения фумарата, чтобы оба продукта поместились в активном центре.[11]
Мутации
Аденилосукцинатлиаза мутанты может иметь значительно сниженную активность вне зависимости от того, находится ли мутация в активном сайте или за его пределами. Вызывающие заболевание мутанты ASL R396C и R396H находятся на входе в активный центр и имеют более низкий VМаксимум чем ASL дикого типа, но мутанты K246E и L311V, которые находятся вдали от активного сайта, также вызывают снижение VМаксимум. ASL-мутант R194C находится вдали от активного сайта, и хотя он поддерживает VМаксимум как и ASL дикого типа, он оказался наименее конформационно стабильным из пяти мутантов. in vitro и по-прежнему вызывает болезнь.[12]
Механизм
Ранее считалось, что механизм действия аденилосукцинатлиазы представляет собой согласованный катализ, при котором водород на β-углероде (по отношению к выходящему азоту) отводится каталитическим основанием, в то время как уходящий азот протонируется атомом азота. каталитическая кислота для удаления E2.[5] Более свежие данные противоречат этой идее и подтверждают, что механизм на самом деле не согласован, но что абстракция происходит первой, а есть промежуточный карбанион вид, который стабилизирован резонансом. Для обеих реакций, катализируемых ASL, сначала происходит депротонирование углерода β до выходящего азота, затем образование и резонансная стабилизация карбаниона, и, наконец, протонирование уходящего азота, которое вызывает разрыв связи C-N.[9] Экспериментальное подтверждение депротонирования, образования карбаниона и лимитирующей скорости стадии протонирования, вызывающей расщепление, означает, что это механизм E1cb. Самые последние данные предполагают, что каталитической кислотой является His171, который ранее считался каталитическим основанием, и что несколько необычно, что это серин в положении 295, действующий как каталитическое основание. Расщепление аденилосукцината до AMP и фумарата является упорядоченным механизмом uni-bi, что означает, что после расщепления фумарат покидает активный сайт раньше, чем это делает AMP.[13]

Роль в болезни
Мутированная аденилосукцинатлиаза (ASL) вызывает у пациентов клиническое заболевание, которое называют дефицит аденилосукцинатлиазы. Это состояние встречается редко и проявляется в различной степени психомоторная отсталость, аутизм, атрофия мышц, и эпилепсия.[14][15] Точная причина заболевания неизвестна, но возможностей недостаточно. пурин нуклеотид синтез для репликация клеток, неисправность пуриновый нуклеотидный цикл и накопление субстратов до токсичных уровней. Было идентифицировано несколько точечных мутаций, связанных с заболеванием, и те, которые гетерозиготный по точечной мутации здоровы, а вот те, кто гомозиготный развиваются клинические проявления заболевания.[16] Число болезнетворных генотипов продолжает расти по мере того, как обнаруживается больше мутаций, и к настоящему времени выявлено тридцать различных точечных мутаций и одна делеция, которые вызывают дефицит аденилосукцинатлиазы.[17]
Когда субстраты ASL (аденилосуцинат и SAICAR) накапливаются из-за дефицита фермента, они дефосфорилированный и превращаются в сукциниладенозин (S-Ado) и сукциниламиноимидазол карбоксимид рибозид (SAICA рибозид).[18] Обычно эти соединения не присутствуют в спинномозговой жидкости или моче, поскольку ASL воздействует на большинство молекул субстрата, прежде чем они смогут накапливаться и фосфорилироваться.[15] В прошлом не существовало хорошего теста на дефицит аденилосукцинатлиазы, что затрудняло диагностику этого редкого заболевания, но недавно был разработан тест для обнаружения SAICA и S-Ado в моче. Тест недорогой и не дал ложноположительных или ложноотрицательных результатов в небольшой выборке исследователей.[19]
Считается, что рибозид SAICA может быть более токсичным соединением, поскольку он обнаруживается в более высоких концентрациях у пациентов с тяжелыми клиническими симптомами, и некоторые исследователи считают, что S-Ado может даже иметь защитное действие. Необходимо провести дополнительные исследования того, что определяет серьезность заболевания, но нестабильность ASL человека в лабораторных условиях была препятствием для этого исследования.[17]
Терапевтические приложения
По мере роста устойчивости к противомалярийным средствам исследователи ищут новые стратегии для воздействия на Плазмодий паразиты, вызывающие малярия, особенно более смертоносные P. falciparum. Некоторые исследователи предложили рассматривать ASL как потенциальную мишень для лекарств, потому что, несмотря на прерывание de novo путь биосинтеза пурина токсичен для хозяина, Плазмодий ASL имеет низкий уровень гомология последовательностей с человеческим ASL, который может сделать любую анти-Плазмодий Препараты ASL достаточно специфичны, чтобы не причинить вред человеку-хозяину.[20]
Рекомендации
- ^ а б c ГРЧ38: Ансамбль выпуск 89: ENSG00000239900 - Ансамбль, Май 2017
- ^ а б c GRCm38: выпуск Ensembl 89: ENSMUSG00000022407 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ «Ссылка на Mouse PubMed:». Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ а б c d е Тот EA, Йейтс TO (февраль 2000 г.). «Структура аденилосукцинатлиазы, фермента с двойной активностью в пути биосинтеза пуринов de novo». Структура. 8 (2): 163–74. Дои:10.1016 / S0969-2126 (00) 00092-7. PMID 10673438.
- ^ "Entrez Gene: аденилосукцинатлиаза". Получено 2012-03-01.
- ^ Селвуд Т., Джаффе Е.К. (март 2012 г.). «Динамические диссоциирующие гомоолигомеры и контроль функции белка». Архивы биохимии и биофизики. 519 (2): 131–43. Дои:10.1016 / j.abb.2011.11.020. ЧВК 3298769. PMID 22182754.
- ^ Spiegel EK, Colman RF, Patterson D (2006). «Недостаток аденилосукцинатлиазы». Молекулярная генетика и метаболизм. 89 (1–2): 19–31. Дои:10.1016 / j.ymgme.2006.04.018. PMID 16839792.
- ^ а б c d Цай М., Ку Дж., Ип П., Колман Р.Ф., Сегалл М.Л., Хауэлл П.Л. (июль 2007 г.). «Комплексы субстрата и продукта аденилосукцинатлиазы Escherichia coli позволяют по-новому взглянуть на ферментативный механизм». Журнал молекулярной биологии. 370 (3): 541–54. Дои:10.1016 / j.jmb.2007.04.052. ЧВК 4113493. PMID 17531264.
- ^ Сивендран С., Колман РФ (июль 2008 г.). «Влияние нового нерасщепляемого аналога субстрата на мутанты дикого типа и сериновые мутанты в сигнатурной последовательности аденилосукцинатлиазы Bacillus subtilis и Homo sapiens». Белковая наука. 17 (7): 1162–74. Дои:10.1110 / пс 034777.108. ЧВК 2442012. PMID 18469177.
- ^ Козлов Г., Нгуен Л., Пирсол Дж., Геринг К. (сентябрь 2009 г.). «Структура фосфат-связанной аденилосукцинатлиазы Escherichia coli идентифицирует His171 как каталитическую кислоту». Acta Crystallographica Раздел F. 65 (Pt 9): 857–61. Дои:10.1107 / S1744309109029674. ЧВК 2795585. PMID 19724117.
- ^ Ariyananda Lde Z, Lee P, Antonopoulos C, Colman RF (июнь 2009 г.). «Биохимический и биофизический анализ пяти мутантов аденилосукцинатлиазы человека, связанных с заболеванием». Биохимия. 48 (23): 5291–302. Дои:10.1021 / bi802321m. ЧВК 2745324. PMID 19405474.
- ^ Булусу В., Сринивасан Б., Бопанна М.П., Баларам Х. (апрель 2009 г.). «Выяснение специфичности субстрата, кинетического и каталитического механизма аденилосукцинатлиазы из Plasmodium falciparum». Biochimica et Biophysica Acta (BBA) - Белки и протеомика. 1794 (4): 642–54. Дои:10.1016 / j.bbapap.2008.11.021. PMID 19111634.
- ^ Маасвинкель-Муйдж, Лаан Л.А., Онкенхаут В., Брауэр О.Ф., Джекен Дж., Поортуис Б.Дж. (август 1997 г.). «Дефицит аденилосукциназы при эпилепсии в раннем младенчестве». Журнал наследственных метаболических заболеваний. 20 (4): 606–7. Дои:10.1023 / А: 1005323512982. PMID 9266401.
- ^ а б Ли П., Колман РФ (февраль 2007 г.). «Экспрессия, очистка и характеристика стабильной рекомбинантной аденилосукцинатлиазы человека». Экспрессия и очистка белков. 51 (2): 227–34. Дои:10.1016 / j.pep.2006.07.023. PMID 16973378.
- ^ Стоун Р.Л., Эйми Дж., Баршоп Б.А., Джекен Дж., Ван ден Берге Дж., Залкин Х., Диксон Дж. Э. (апрель 1992 г.). «Мутация аденилосукцинатлиазы, связанная с умственной отсталостью и особенностями аутизма». Природа Генетика. 1 (1): 59–63. Дои:10.1038 / ng0492-59. PMID 1302001.
- ^ а б Паленчар Дж. Б., Крокко Дж. М., Колман РФ (август 2003 г.). «Характеристика мутантных аденилосукцинатлиаз Bacillus subtilis, соответствующих тяжелой недостаточности аденилосукцинатлиазы человека». Белковая наука. 12 (8): 1694–705. Дои:10.1110 / л.с. 0303903. ЧВК 2323956. PMID 12876319.
- ^ Jaeken J, Van den Berghe G (ноябрь 1984 г.). «Инфантильный аутистический синдром, характеризующийся наличием сукцинилпуринов в жидкостях организма». Ланцет. 2 (8411): 1058–61. Дои:10.1016 / с0140-6736 (84) 91505-8. PMID 6150139.
- ^ Мэддокс Дж., Рид Т. (январь 1989 г.). «Анализ мочи на дефицит аденилосукциназы у детей-аутистов». Ланцет. 1 (8630): 158–9. Дои:10.1016 / S0140-6736 (89) 91172-0. PMID 2563072.
- ^ Маршалл В.М., Коппель Р.Л. (сентябрь 1997 г.). «Характеристика гена, кодирующего аденилосукцинатлиазу Plasmodium falciparum». Молекулярная и биохимическая паразитология. 88 (1–2): 237–41. Дои:10.1016 / S0166-6851 (97) 00054-6. PMID 9274883.
дальнейшее чтение
- Marie S, Cuppens H, Heuterspreute M, Jaspers M, Tola EZ, Gu XX, Legius E, Vincent MF, Jaeken J, Cassiman JJ, Van den Berghe G (1999). «Мутационный анализ дефицита аденилосукцинатлиазы: восемь новых мутаций в пересмотренной полной кодирующей последовательности ADSL». Человеческая мутация. 13 (3): 197–202. Дои:10.1002 / (SICI) 1098-1004 (1999) 13: 3 <197 :: AID-HUMU3> 3.0.CO; 2-D. PMID 10090474.
- Kmoch S, Hartmannová H, Stibůrková B, Krijt J, Zikánová M, Sebesta I. (июнь 2000 г.). «Аденилосукцинатлиаза человека (ADSL), клонирование и характеристика полноразмерной кДНК и ее изоформы, генная структура и молекулярные основы дефицита ADSL у шести пациентов». Молекулярная генетика человека. 9 (10): 1501–13. Дои:10.1093 / hmg / 9.10.1501. PMID 10888601.
- Race V, Marie S, Vincent MF, Van den Berghe G (сентябрь 2000 г.). «Клинические, биохимические и молекулярно-генетические корреляции при дефиците аденилосукцинатлиазы». Молекулярная генетика человека. 9 (14): 2159–65. Дои:10.1093 / hmg / 9.14.2159. PMID 10958654.
- Табуччи А., Карлуччи Ф., Рози Ф., Герранти Р., Маринелло Е. (июнь 2001 г.). «Определение, активность и биологическая роль аденилосукцинатлиазы в клетках крови». Биомедицина и фармакотерапия. 55 (5): 277–83. Дои:10.1016 / s0753-3322 (01) 00061-0. PMID 11428554.
- Marie S, Race V, Nassogne MC, Vincent MF, Van den Berghe G (июль 2002 г.). «Мутация сайта связывания ядерного респираторного фактора 2 в 5'-нетранслируемой области гена ADSL у трех пациентов с дефицитом аденилосукцинатлиазы». Американский журнал генетики человека. 71 (1): 14–21. Дои:10.1086/341036. ЧВК 384970. PMID 12016589.
- Кастро М., Перес-Серда С., Меринеро Б., Гарсиа М.Дж., Бернар Дж., Гил Нагель А., Торрес Дж., Бермудес М., Гаравито П., Мари С., Винсент Ф., Ван ден Берге Г., Угарте М. (август 2002 г.). «Скрининг дефицита аденилосукцинатлиазы: клинические, биохимические и молекулярные данные у четырех пациентов». Нейропедиатрия. 33 (4): 186–9. Дои:10.1055 / с-2002-34493. PMID 12368987.
- Паленчар Дж. Б., Колман РФ (февраль 2003 г.). «Характеристика мутантной аденилосукцинатлиазы Bacillus subtilis, эквивалентной мутантному ферменту, обнаруженному при дефиците аденилосукцинатлиазы человека: аспарагин 276 играет важную структурную роль». Биохимия. 42 (7): 1831–41. Дои:10.1021 / bi020640 +. PMID 12590570.
- Эдери П., Шабрие С., Себальо-Пико I, Мари С., Винсент М.Ф., Тардье М. (июль 2003 г.). «Внутрисемейная изменчивость фенотипического выражения недостаточности аденилосукцинатлиазы: отчет о трех пациентах». Американский журнал медицинской генетики, часть A. 120А (2): 185–90. Дои:10.1002 / ajmg.a.20176. PMID 12833398.
- Стоун Р.Л., Эйми Дж., Баршоп Б.А., Джекен Дж., Ван ден Берге Дж., Залкин Х., Диксон Дж. Э. (апрель 1992 г.). «Мутация аденилосукцинатлиазы, связанная с умственной отсталостью и особенностями аутизма». Природа Генетика. 1 (1): 59–63. Дои:10.1038 / ng0492-59. PMID 1302001.
- Сивендран С., Паттерсон Д., Шпигель Э, МакГаун И., Коули Д., Колман Р.Ф. (декабрь 2004 г.). «Две новые мутантные аденилосукцинатлиазы человека (ASL), связанные с аутизмом, и характеристика эквивалентного мутантного ASL Bacillus subtilis». Журнал биологической химии. 279 (51): 53789–97. Дои:10.1074 / jbc.M409974200. PMID 15471876.
внешняя ссылка
- Аденилосукцинат + лиаза в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- Человек ADSL расположение генома и ADSL страница сведений о генах в Браузер генома UCSC.
- Человек ASL расположение генома и ASL страница сведений о генах в Браузер генома UCSC.