Ацетил-КоА карбоксилаза - Acetyl-CoA carboxylase
| Ацетил-КоА карбоксилаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Номер ЕС | 6.4.1.2 | ||||||||
| Количество CAS | 9023-93-2 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
| Ацетил-КоА карбоксилаза альфа | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Символ | ACACA | ||||||
| Альт. символы | ACAC, ACC1, ACCA | ||||||
| Ген NCBI | 31 | ||||||
| HGNC | 84 | ||||||
| OMIM | 601557 | ||||||
| RefSeq | NM_198839 | ||||||
| UniProt | Q13085 | ||||||
| Прочие данные | |||||||
| Номер ЕС | 6.4.1.2 | ||||||
| Locus | Chr. 17 q21 | ||||||
| |||||||
| Ацетил-КоА карбоксилаза бета | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Символ | ACACB | ||||||
| Альт. символы | ACC2, ACCB | ||||||
| Ген NCBI | 32 | ||||||
| HGNC | 85 | ||||||
| OMIM | 200350 | ||||||
| RefSeq | NM_001093 | ||||||
| UniProt | O00763 | ||||||
| Прочие данные | |||||||
| Номер ЕС | 6.4.1.2 | ||||||
| Locus | Chr. 12 q24.1 | ||||||
| |||||||
Ацетил-КоА карбоксилаза (АКК) это биотин -зависимый фермент что катализирует необратимые карбоксилирование из ацетил-КоА производить малонил-КоА благодаря двум его каталитическим действиям, биотин карбоксилаза (BC) и карбоксилтрансфераза (КТ). АЦЦ - это мультисубъединичный фермент в большинстве прокариоты и в хлоропласты большинства растений и водорослей, в то время как это большой мультидоменный фермент в эндоплазматический ретикулум из большинства эукариоты. Наиболее важной функцией ACC является обеспечение субстрата малонил-КоА для биосинтез жирных кислот.[1] Активностью ACC можно управлять на уровне транскрипции, а также с помощью модуляторов малых молекул и ковалентная модификация. Геном человека содержит гены двух разных АСС.[2]—ACACA[3] и ACACB.[4]
Структура
Прокариоты и растения имеют мультисубъединичные АСС, состоящие из нескольких полипептидов. Активность биотинкарбоксилазы (BC), белок-носитель карбоксила биотина (BCCP) и активность карбоксилтрансферазы (CT) каждая содержится в разных субъединицах. Стехиометрия этих субъединиц в ACC холоэнзим отличается среди организмов.[1] Люди и большинство эукариоты развили ACC с каталитическими доменами CT и BC и доменами BCCP на единственном полипептиде. Большинство растений также имеют эту гомомерную форму в цитозоле.[5] Функциональные области ACC, начиная с N-конец к C-конец представляют собой биотинкарбоксилазу (BC), связывание биотина (BB), карбоксилтрансферазу (CT) и АТФ-связывание (AB). AB находится внутри BC. Биотин ковалентно присоединен через амидную связь к длинному боковая цепь лизина находятся в BB. Поскольку BB находится между BC и CT областями, биотин может легко перемещаться в оба активных центра, где это необходимо.
У млекопитающих, у которых экспрессируются две изоформы ACC, основным структурным различием между этими изоформами является удлиненный N-конец ACC2, содержащий митохондриальная нацеливающая последовательность.[1]
- Кристаллографические структуры Кишечная палочка ацетил-КоА карбоксилаза

Субъединица биотинкарбоксилазы Кишечная палочка ацетил-КоА карбоксилаза

Биотин карбоксильная белковая субъединица Кишечная палочка ацетил-КоА карбоксилаза

Субъединица карбоксилтрансферазы Кишечная палочка ацетил-КоА карбоксилаза


Гены
Полипептиды, составляющие мульти-субъединичные АСС прокариоты и растения кодируются разными генами. В кишечная палочка, accA кодирует альфа-субъединицу ацетил-КоА карбоксилазы,[6] и accD кодирует свою бета-субъединицу.[7]
Механизм
Общая реакция ACAC (A, B) протекает по двухступенчатому механизму.[8] Первая реакция осуществляется BC и включает АТФ-зависимое карбоксилирование биотин с участием бикарбонат служащий источником CO2. Карбоксильная группа переносится из биотин к ацетил-КоА формировать малонил-КоА во второй реакции, катализируемой ХТ.
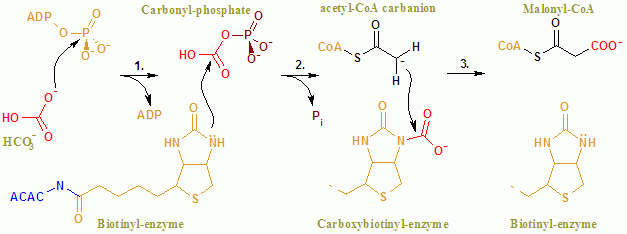
в активный сайт, реакция протекает при обширном взаимодействии остатков Glu296 и положительно заряженных Arg338 и Arg292 с субстратами.[9] Два мг2+ координируются фосфатными группами на АТФ, и необходимы для связывания АТФ с ферментом. Бикарбонат депротонированный Glu296, хотя в растворе этот перенос протона маловероятен, поскольку pKa бикарбоната составляет 10,3. Фермент, по-видимому, манипулирует pKa, чтобы облегчить депротонирование бикарбоната. PKa бикарбоната снижается за счет его взаимодействия с положительно заряженными боковыми цепями Arg338 и Arg292. Кроме того, Glu296 взаимодействует с боковой цепью Glu211, взаимодействие, которое, как было показано, вызывает увеличение видимого pKa. После депротонирования бикарбоната кислород бикарбоната действует как нуклеофил и атакует гамма-фосфат на АТФ. Карбоксифосфатный промежуточный продукт быстро разлагается до CO.2 и ПО43−. ПО43− депротонирует биотин, создавая енолят, стабилизированный Arg338, который впоследствии атакует CO2 в результате производства карбоксибиотина.[9] Карбоксибиотин перемещается в активный центр карбоксилтрансферазы (СТ), где карбоксильная группа переносится на ацетил-КоА. В отличие от домена BC, о механизме реакции CT известно немного. Предлагаемый механизм - выброс CO2 от биотина, который впоследствии отрывает протон от метильной группы от ацетил-СоА-карбоксилазы. Результирующий энолировать атакует CO2 с образованием малонил-КоА. В конкурирующем механизме протон абстракция связана с атакой ацетил-КоА.
Функция
Функция ACC - регулировать метаболизм жирных кислот. Когда фермент активен, образуется продукт малонил-КоА, который является строительным блоком для новых жирных кислот и может ингибировать перенос жирной ацильной группы от ацил-КоА к карнитин с участием карнитинацилтрансфераза, что подавляет бета-окисление жирных кислот в митохондрии.
В млекопитающие, два основных изоформы ACC экспрессируются, ACC1 и ACC2, которые различаются как распределением в тканях, так и функцией. ACC1 находится в цитоплазма всех клеток, но обогащен липогенной тканью, такой как жировой ткани и кормящие молочные железы, где важен синтез жирных кислот.[10] В окислительных тканях, таких как скелетная мышца и сердце, соотношение выраженного ACC2 выше. ACC1 и ACC2 высоко экспрессируются в печень где важны как окисление, так и синтез жирных кислот.[11] Различия в распределении тканей указывают на то, что ACC1 поддерживает регуляцию синтез жирных кислот тогда как ACC2 в основном регулирует окисление жирных кислот (бета-окисление).
Регулирование

Регулирование ACC млекопитающих является сложным, чтобы контролировать два различных пула малонил-КоА, которые управляют либо ингибированием бета-окисления, либо активацией биосинтеза липидов.[12]
ACC1 и ACC2 млекопитающих регулируются транскрипционно множеством промоутеры которые опосредуют изобилие ACC в ответ на состояние питания клеток. Активация экспрессии генов через разные промоторы приводит к альтернативное сращивание; однако физиологическое значение конкретного АСС изоферменты остается неясным.[11] Чувствительность к статусу питания является результатом контроля этих промоторов с помощью факторы транскрипции такие как белок 1, связывающий регуляторный элемент стерола, контролируется инсулином на уровне транскрипции, и ЧРЭБП, выражение которого увеличивается с увеличением углеводы диеты.[13][14]
Через петлю прямой связи, цитрат аллостерически активирует АСС.[15] Цитрат может увеличить АСС полимеризация для повышения ферментативной активности; однако неясно, является ли полимеризация основным механизмом цитратного увеличения активности АЦЦ или полимеризация является артефактом экспериментов in vitro. Другие аллостерические активаторы включают: глутамат и другие дикарбоновые кислоты.[16] Длинноцепочечные и короткоцепочечные жирные ацил-КоА являются ингибиторами АСС с отрицательной обратной связью.[17]
Фосфорилирование может произойти, когда гормоны глюкагон или адреналин связываться с поверхностью клетки рецепторы, но основная причина фосфорилирования связана с повышением уровней АМФ при низком энергетическом статусе клетки, что приводит к активации АМФ-активированная протеинкиназа (АМПК). АМПК является основным киназа регулятор ACC, способный фосфорилировать ряд остатков серина на обеих изоформах ACC.[18] На ACC1 AMPK фосфорилирует Ser79, Ser1200 и Ser1215. Протеинкиназа А также обладает способностью фосфорилировать ACC с гораздо большей способностью фосфорилировать ACC2, чем ACC1. Однако физиологическое значение протеинкиназы A в регуляции ACC в настоящее время неизвестно. Исследователи предполагают, что существуют другие киназы ACC, важные для его регуляции, поскольку существует множество других возможных сайтов фосфорилирования на ACC.[19]
Когда инсулин связывается со своими рецепторами на клеточная мембрана, он активирует фермент фосфатазу, называемый протеинфосфатаза 2А (PP2A) для дефосфорилирования фермента; тем самым снимая тормозящий эффект. Кроме того, инсулин индуцирует фосфодиэстеразу, которая снижает уровень цАМФ в клетке, таким образом ингибируя PKA, а также напрямую ингибирует AMPK.[нужна цитата ]
Этот белок может использовать морфеин модель аллостерическая регуляция.[20]
Клинические последствия
На стыке путей синтеза и окисления липидов ACC представляет множество клинических возможностей для производства новых антибиотики и разработка новых методов лечения сахарный диабет, ожирение, и другие проявления метаболический синдром.[21] Исследователи стремятся использовать структурные различия между бактериальными и человеческими ACC для создания антибиотиков, специфичных для бактериальных ACC, чтобы минимизировать побочные эффекты для пациентов. Многообещающие результаты полезности ингибитора АСС включают открытие, что у мышей без экспрессии АСС2 наблюдается непрерывное окисление жирных кислот, сниженная масса тела и сниженная масса тела, несмотря на увеличение потребления пищи. Эти мыши также защищены от диабета.[12] Недостаток ACC1 у мутантных мышей летален уже на эмбриональной стадии. Однако неизвестно, должны ли препараты, нацеленные на ACC у людей, быть специфичными для ACC2.[22]
Фирсокостат (ранее GS-976, ND-630, NDI-010976) является мощным аллостерическим ингибитором ACC, действующим на BC-домен ACC.[23] Фирсокостат находится в разработке в 2019 г. (Фаза II)[24] фармацевтической компанией Галаад в составе комбинированного лечения неалкогольный стеатогепатит (НАСГ), который считается все более частой причиной печеночной недостаточности.[25]
Кроме того, селективные для растений ингибиторы АЦК широко используются в качестве гербициды,[26] что предполагает клиническое применение против Apicomplexa паразиты, которые полагаются на изоформу АЦК растительного происхождения,[27] в том числе малярия.
Смотрите также
использованная литература
- ^ а б c Тонг Л. (август 2005 г.). «Ацетил-кофермент карбоксилаза: важнейший метаболический фермент и привлекательная цель для открытия новых лекарств». Клеточные и молекулярные науки о жизни. 62 (16): 1784–803. Дои:10.1007 / s00018-005-5121-4. PMID 15968460. S2CID 1131957.
- ^ Браунси Р. У., Жанде Р., Бун А. Н. (ноябрь 1997 г.). «Изоформы ацетил-КоА карбоксилазы: структуры, регуляторные свойства и метаболические функции». Сделки Биохимического Общества. 25 (4): 1232–8. Дои:10.1042 / bst0251232. PMID 9449982.
- ^ Абу-Эльхейга Л., Джаякумар А., Балдини А., Чирала С.С., Вакил С.Дж. (апрель 1995 г.). «Ацетил-КоА-карбоксилаза человека: характеристика, молекулярное клонирование и доказательства двух изоформ». Труды Национальной академии наук Соединенных Штатов Америки. 92 (9): 4011–5. Bibcode:1995PNAS ... 92.4011A. Дои:10.1073 / пнас.92.9.4011. ЧВК 42092. PMID 7732023.
- ^ Видмер Дж., Фассихи К.С., Шлихтер С.К., Уиллер К.С., Crute BE, King N, Nutile-McMenemy N, Noll WW, Daniel S, Ha J, Kim KH, Witters LA (июнь 1996 г.). «Идентификация второго гена ацетил-КоА-карбоксилазы человека». Биохимический журнал. 316 (3): 915–22. Дои:10.1042 / bj3160915. ЧВК 1217437. PMID 8670171.
- ^ Сасаки Ю., Нагано И. (июнь 2004 г.). «Ацетил-КоА-карбоксилаза растений: структура, биосинтез, регуляция и генные манипуляции для селекции растений». Биология, биотехнология и биохимия. 68 (6): 1175–84. Дои:10.1271 / bbb.68.1175. PMID 15215578. S2CID 41506311.
- ^ "accA, альфа-субъединица ацетил-КоА карбоксилазы (кишечная палочка ул. К-12 подл. MG1655) ". Ген NCBI. Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ "accD, бета-субъединица ацетил-КоА карбоксилазы (кишечная палочка ул. К-12 подл. MG1655) ". Ген NCBI. Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ Lee CK, Cheong HK, Ryu KS, Lee JI, Lee W, Jeon YH, Cheong C (август 2008 г.). «Биотиноильный домен ацетил-CoA-карбоксилазы человека: структурное понимание механизма переноса карбоксила». Белки. 72 (2): 613–24. Дои:10.1002 / prot.21952. PMID 18247344. S2CID 24548083.
- ^ а б Чжоу CY, Yu LP, Tong L (апрель 2009 г.). «Кристаллическая структура биотинкарбоксилазы в комплексе с субстратами и последствия для ее каталитического механизма». Журнал биологической химии. 284 (17): 11690–7. Дои:10.1074 / jbc.M805783200. ЧВК 2670172. PMID 19213731.
- ^ Ким Т.С., Лихи П., Фрик ХК (август 1996 г.). «Использование промотора определяет тканеспецифическую чувствительность гена ацетил-КоА-карбоксилазы крысы». Сообщения о биохимических и биофизических исследованиях. 225 (2): 647–53. Дои:10.1006 / bbrc.1996.1224. PMID 8753813.
- ^ а б Барбер М.С., Price NT, Travers MT (март 2005 г.). «Структура и регуляция генов ацетил-CoA карбоксилазы многоклеточных». Biochimica et Biophysica Acta (BBA) - Молекулярная и клеточная биология липидов. 1733 (1): 1–28. Дои:10.1016 / j.bbalip.2004.12.001. PMID 15749055.
- ^ а б Абу-Эльхейга Л., Мацук М.М., Або-Хашема К.А., Вакил С.Дж. (март 2001 г.). «Непрерывное окисление жирных кислот и снижение накопления жира у мышей, лишенных ацетил-КоА-карбоксилазы 2». Наука. 291 (5513): 2613–6. Bibcode:2001Научный ... 291.2613A. Дои:10.1126 / science.1056843. PMID 11283375. S2CID 748630.
- ^ Филд FJ, Born E, Murthy S, Mathur SN (декабрь 2002 г.). «Полиненасыщенные жирные кислоты снижают экспрессию белка-1, связывающего регуляторный элемент стерола, в клетках CaCo-2: влияние на синтез жирных кислот и транспорт триацилглицерина». Биохимический журнал. 368 (Pt 3): 855–64. Дои:10.1042 / BJ20020731. ЧВК 1223029. PMID 12213084.
- ^ Исии С., Иидзука К., Миллер BC, Уеда К. (ноябрь 2004 г.). «Белок, связывающий элемент углеводного ответа, напрямую способствует транскрипции гена липогенного фермента». Труды Национальной академии наук Соединенных Штатов Америки. 101 (44): 15597–602. Bibcode:2004PNAS..10115597I. Дои:10.1073 / pnas.0405238101. ЧВК 524841. PMID 15496471.
- ^ Мартин Д. Б., Вагелос PR (июнь 1962 г.). «Механизм регуляции цикла трикарбоновых кислот синтеза жирных кислот». Журнал биологической химии. 237: 1787–92. PMID 14470343.
- ^ Бун А.Н., Чан А., Кульпа Дж. Э., Браунси Р. В. (апрель 2000 г.). «Бимодальная активация ацетил-КоА карбоксилазы глутаматом». Журнал биологической химии. 275 (15): 10819–25. Дои:10.1074 / jbc.275.15.10819. PMID 10753875.
- ^ Фаэргеман Н. Дж., Кнудсен Дж. (Апрель 1997 г.). «Роль длинноцепочечных сложных эфиров ацил-КоА с длинной цепью в регуляции метаболизма и клеточной сигнализации». Биохимический журнал. 323 (Чт 1): 1–12. Дои:10.1042 / bj3230001. ЧВК 1218279. PMID 9173866.
- ^ Park SH, Gammon SR, Knippers JD, Paulsen SR, Rubink DS, Winder WW (июнь 2002 г.). «Отношения фосфорилирования-активности AMPK и ацетил-КоА карбоксилазы в мышцах». Журнал прикладной физиологии. 92 (6): 2475–82. Дои:10.1152 / japplphysiol.00071.2002. PMID 12015362.
- ^ Браунси Р. У., Бун А. Н., Эллиотт Дж. Э., Кульпа Дж. Э., Ли В. М. (апрель 2006 г.). «Регулирование ацетил-КоА карбоксилазы». Сделки Биохимического Общества. 34 (Чт 2): 223–7. Дои:10.1042 / BST20060223. PMID 16545081.
- ^ Селвуд Т., Джаффе Е.К. (март 2012 г.). «Динамические диссоциирующие гомоолигомеры и контроль функции белка». Архивы биохимии и биофизики. 519 (2): 131–43. Дои:10.1016 / j.abb.2011.11.020. ЧВК 3298769. PMID 22182754.
- ^ Корбетт Дж. В., Харвуд Дж. Х. (ноябрь 2007 г.). «Ингибиторы ацетил-КоА карбоксилазы млекопитающих». Последние патенты на открытие сердечно-сосудистых лекарств. 2 (3): 162–80. Дои:10.2174/157489007782418928. PMID 18221116.
- ^ Абу-Эльхейга Л., Мацук М.М., Кордари П., О В., Шайкенов Т., Гу З., Вакил С.Дж. (август 2005 г.). «Мутантные мыши, лишенные ацетил-КоА карбоксилазы 1, эмбрионально летальны». Труды Национальной академии наук Соединенных Штатов Америки. 102 (34): 12011–6. Bibcode:2005PNAS..10212011A. Дои:10.1073 / pnas.0505714102. ЧВК 1189351. PMID 16103361.
- ^ Харриман Дж., Гринвуд Дж., Бхат С., Хуанг Х, Ван Р., Пол Д., Тонг Л., Саха А. К., Вестлин В. Ф., Капеллер Р., Харвуд Х. Дж. (Март 2016 г.). «Ингибирование ацетил-КоА-карбоксилазы ND-630 снижает стеатоз печени, улучшает чувствительность к инсулину и модулирует дислипидемию у крыс». Труды Национальной академии наук Соединенных Штатов Америки. 113 (13): E1796–805. Bibcode:2016PNAS..113E1796H. Дои:10.1073 / pnas.1520686113. ЧВК 4822632. PMID 26976583.
- ^ Тонг А (11 апреля 2019 г.). «Gilead вселяет надежду на коктейль NASH, взглянув на положительные данные, подтверждающие правильность концепции». Новости конечных точек.
- ^ Lucas C, Lucas G, Lucas N, Krzowska-Firych J, Tomasiewicz K (сентябрь 2018 г.). «Систематический обзор настоящего и будущего неалкогольной жировой болезни печени». Клиническая и экспериментальная гепатология. 4 (3): 165–174. Дои:10.5114 / ceh.2018.78120. ЧВК 6185929. PMID 30324141.
- ^ Аль-Хатиб К. «Ингибиторы ацетил-КоА-карбоксилазы (АССазы)». Симптомы гербицидов. Отдел сельского хозяйства и природных ресурсов Калифорнийского университета в Дэвисе.
- ^ Зютер Э., Джонсон Дж. Дж., Хаселкорн Р., МакЛеод Р., Горницки П. (ноябрь 1999 г.). «Рост Toxoplasma gondii подавляется арилоксифеноксипропионатными гербицидами, нацеленными на ацетил-КоА-карбоксилазу». Труды Национальной академии наук Соединенных Штатов Америки. 96 (23): 13387–92. Bibcode:1999PNAS ... 9613387Z. Дои:10.1073 / пнас.96.23.13387. ЧВК 23957. PMID 10557330.
дальнейшее чтение
- Воет Д., Воет Дж. Г. (2004). Биохимия (3-е изд.). Вайли. ISBN 978-0-471-19350-0.
- Бьюкенен BB, Gruissem W, Jones RL, ред. (2000). Биохимия и молекулярная биология растений. Американское общество физиологов растений. ISBN 978-0-943088-37-2.
- Леверт К.Л., Уолдроп Г.Л., Стивенс Дж.М. (май 2002 г.). «Аналог биотина ингибирует активность ацетил-КоА-карбоксилазы и адипогенез». Журнал биологической химии. 277 (19): 16347–50. Дои:10.1074 / jbc.C200113200. PMID 11907024.