Пантотенаткиназа - Pantothenate kinase
| Пантотенаткиназа | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Номер ЕС | 2.7.1.33 | ||||||||
| Количество CAS | 9026-48-6 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
Пантотенаткиназа (ЕС 2.7.1.33, PanK; CoaA) является первым фермент в Коэнзим А (КоА) биосинтетический путь. Фосфорилирует пантотенат (витамин B5) с образованием 4'-фосфопантотената за счет молекулы аденозинтрифосфат (АТФ ). Это лимитирующий этап биосинтеза КоА.[1][2]
![[2]](http://upload.wikimedia.org/wikipedia/commons/3/34/Mechanism_os_pantothenate_kinase.png)
CoA является необходимым кофактор во всех живых организмах. Он выступает в качестве основного ацильная группа носитель во многих важных клеточных процессах, таких как цикл лимонной кислоты (цикл трикарбоновых кислот ) и жирная кислота метаболизм. Следовательно, пантотенаткиназа является ключевым регуляторным ферментом в пути биосинтеза КоА.[3]
Типы
Было идентифицировано три различных типа PanK - PanK-I (обнаружен в бактериях), PanK-II (в основном обнаружен у эукариот, но также и в Стафилококки ) и PanK-III, также известный как CoaX (обнаружен в бактериях). Эукариотические ферменты PanK-II часто встречаются в виде разных изоформ, таких как PanK1, PanK2, PanK3 и PanK4. У человека несколько изоформ PanK экспрессируются четырьмя генами. Ген PANK1 кодирует формы PanK1α и PanK1β, а PANK2 и PANK3 кодируют PanK2 и PanK3 соответственно.[4]
Структура
ПанК-II

PanK-II состоит из двух белковые домены, как показано на рисунке 1. Каждый домен A и A 'имеет богатую глицином петлю (последовательность GXXXXGKS; Петля P ), что характерно для нуклеотид -участок связывания; это где АТФ предполагается связать.[5] расположен между остатками 95 и 102 в домене A
Два АТФ отображение сайтов привязки совместное поведение. Интерфейс димеризации состоит из двух длинных спиралей, по одной от каждого мономера, которые взаимодействуют друг с другом. В C-терминал концы спиралей скрепляются Ван-дер-Ваальсовы взаимодействия между валин и метионин остатки каждого мономера. Середина спиралей прикреплена водородными связями между аспарагин остатки. На N-концевой В конце каждая спираль расширяется и образует четырехспиральный пучок с двумя более короткими спиралями. Этот пучок состоит из гидрофобного ядра, образованного неполярными остатками, которые используют силы Ван-дер-Ваальса для дальнейшей стабилизации димера.[4]
В активном центре пантотенат ориентирован водородными связями между пантотенатом и боковыми цепями остатков аспартата, тирозина, гистидина, тирозина и аспарагина.[6] Остатки аспарагина, гистидина и аргинина участвуют в катализе.
Изоформы PanK-II человека PanK1α, PanK1β, PanK2 и PanK3 имеют общее высокогомологичное каталитическое ядро из примерно 355 остатков.[4] PanK1α и PanK1β оба кодируются PANK1 ген и имеют одинаковый каталитический домен из 363 аминокислот, кодируемый экзоны 2-7. Транскрипт PanK1α начинается с экзона 1α, который кодирует регуляторный домен из 184 остатков в N-конец. Эта область позволяет подавлять обратную связь свободным CoA и ацил-КоА и регулирование ацетил-КоА и малонил-КоА. С другой стороны, транскрипт PanK1β начинается с экзона 1β, который дает N-конец из 10 остатков, который не включает регуляторный домен обратной связи.[7]
ПанК-III

PanK-III также содержит два белковые домены, а ключевые каталитические остатки PanK-II сохранены. Мономерные звенья PanK-II и PanK-III практически идентичны, но имеют совершенно разные димерные сборки. Исследование между структурами Золотистый стафилококк тип II и Синегнойная палочка тип III демонстрирует, что мономер PanK-II имеет область петли, которая отсутствует в мономере PanK-III, а мономер PanK-III имеет область петли, которая отсутствует в мономере PanK-II.[8] Это незначительное изменение имеет решающее различие на границе димеризации, в которой спирали димера PanK-II наматываются друг на друга, а спирали димера PanK-III взаимодействуют под углом 70 ° (Рис. 2).[9]
В результате этой разницы в интерфейсе димеризации между PanK-II и PanK-III конформации сайтов связывания субстрата для АТФ и пантотенат также различны.[10][11]
Каталитический механизм
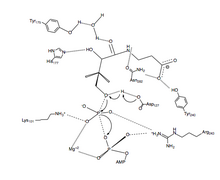
ПанК-II
Предлагаемый механизм реакции переноса фосфорила PanK-II представляет собой согласованный механизм с диссоциативным переходное состояние.
Во-первых, АТФ связывается в канавке для связывания, образованной остатками Петля P и близлежащие остатки. Здесь консервированные лизин (Лиз-101 ) - ключевой остаток, необходимый для АТФ привязка.[12][13] Кроме того, боковые цепи остатков Лиз-101, Сер-102, Glu-199, и Арг-243 ориентировать нуклеотид в паз для обвязки. В пантотенат связан и ориентирован путем формирования взаимодействия водородных связей с остатками Asp-127, Тир-240, Асн-282, Тир-175, и Его-177.[6] Когда оба АТФ и пантотенат связаны, Asp-127 депротонирует C1 гидроксил группа пантотенат. Кислород из пантотенат затем атакует γ-фосфат связанного АТФ. Здесь стабилизация заряда β- и γ-фосфатных групп достигается за счет Арг-243, Лиз-101, и согласованный Mg2+ ион.[14] В этом согласованном механизме планарный фосфоран γ-фосфата передается в линию атакующему кислороду пантотенат.[6] Наконец, 4'-фосфопантотенат диссоциирует от PanK с последующим ADP.
Регулирование пантотенаткиназы
ПанК-II
Регулирование пантотенаткиназы имеет важное значение для контроля внутриклеточного CoA концентрация.[15] Пантотенаткиназа регулируется посредством ингибирования обратной связи посредством CoA и его тиоэфиры (т.е. ацетил-КоА, малонил-КоА ).[16] CoA подавляет активность PanK за счет конкурентно обязательный к АТФ сайт связывания и предотвращение связывания АТФ с Lys-101.[12][13]
Несмотря на то что CoA связывается на том же сайте, что и АТФ, они связываются в разных ориентациях, и их аденин фрагменты взаимодействуют с фермент с неперекрывающимися множествами вычетов. Гис-177, Phe-247, и Арг-106 необходимы для CoA признание, но не для АТФ, а пока Асн-43 и His-307 взаимодействовать с аденин основа АТФ, Его-177 и Phe-247 взаимодействовать с аденин основа CoA.[14] Обе молекулы используют Лиз-101 для нейтрализации заряда соответствующих фосфодиэфиров.
Неэтерифицированный CoA имеет более сильное ингибирование, чем его тиоэфиры. Этот феномен лучше всего объясняется плотной посадкой тиол группа с окружающими ароматическими остатками, Phe-244, Phe-259, Тир-262, и Phe-252. Свободный CoA имеет оптимальную посадку, но когда ацильная группа прикреплен к CoA, стерические препятствия затрудняют взаимодействие тиоэфира с Phe-252. Таким образом, ингибирование тиоэфирами менее эффективно, чем ингибирование неэтерифицированными CoA.[14]
ПанК-III
Вышеуказанное положение соответствует PanK-II. PanK-III устойчив к подавлению обратной связи.[8][10][11]
Гены
В людях:
Ген PANK2 кодирует PanK2, который регулирует образование КоА в митохондрии, центры производства энергии клетки.[17] Мутация PANK2 является причиной Нейродегенерация, связанная с пантотенаткиназой, ранее называвшийся синдромом Халлервордена-Шпатца. Это редкое заболевание проявляется глубокой дистонией, спастичностью и часто приводит к летальному исходу.
использованная литература
- ^ Робишоу Дж. Д., Беркич Д., Нили Дж. Р. (сентябрь 1982 г.). «Ограничение скорости и контроль синтеза кофермента А в сердечной мышце». Журнал биологической химии. 257 (18): 10967–72. PMID 7107640.
- ^ а б Ян К., Эйобо Ю., Брэнд Л.А., Мартыновски Д., Томчик Д., Штраус Е., Чжан Х. (август 2006 г.). «Кристаллическая структура пантотенаткиназы типа III: понимание механизма важного биосинтетического фермента кофермента А, повсеместно распространенного в бактериях». Журнал бактериологии. 188 (15): 5532–40. Дои:10.1128 / JB.00469-06. ЧВК 1540032. PMID 16855243.
- ^ Леонарди Р., Чжан Ю.М., Rock CO, Jackowski S (2005-03-01). «Коэнзим А: снова в действии». Прогресс в исследованиях липидов. 44 (2–3): 125–53. Дои:10.1016 / j.plipres.2005.04.001. PMID 15893380.
- ^ а б c Хонг Б.С., Сенистерра Г., Рабех В.М., Ведади М., Леонарди Р., Чжан Ю.М., Рок СО, Яковски С., Парк Х.В. (сентябрь 2007 г.). «Кристаллические структуры пантотенаткиназ человека. Понимание аллостерической регуляции и мутаций, связанных с нейродегенеративным расстройством». Журнал биологической химии. 282 (38): 27984–93. Дои:10.1074 / jbc.M701915200. PMID 17631502.
- ^ Сарасте М., Сиббальд П.Р., Виттинггофер А (ноябрь 1990 г.). «Р-петля - общий мотив в АТФ- и ГТФ-связывающих белках». Тенденции в биохимических науках. 15 (11): 430–4. Дои:10.1016 / 0968-0004 (90) 90281-Ф. PMID 2126155.
- ^ а б c Айви Р.А., Чжан Ю.М., Вирга К.Г., Хевенер К., Ли Р.Э., Рок СО, Яковски С., Парк Х.В. (август 2004 г.). «Структура тройного комплекса пантотенаткиназа.АДФ.пантотенат выявляет взаимосвязь между сайтами связывания субстрата, аллостерического регулятора и антиметаболитов». Журнал биологической химии. 279 (34): 35622–9. Дои:10.1074 / jbc.M403152200. PMID 15136582.
- ^ Rock CO, Карим MA, Чжан YM, Jackowski S (2002). «Ген мышиной пантотенаткиназы (Pank1) кодирует два дифференциально регулируемых изофермента пантотенаткиназы». Ген. 291 (1–2): 35–43. Дои:10.1016 / S0378-1119 (02) 00564-4. PMID 12095677.
- ^ а б Ян К., Штраус Э, Уэрта С., Чжан Х (февраль 2008 г.). «Структурная основа связывания субстрата и каталитический механизм пантотенаткиназы III типа». Биохимия. 47 (5): 1369–80. Дои:10.1021 / bi7018578. PMID 18186650.
- ^ Хонг Б.С., Юн М.К., Чжан Ю.М., Чохнан С., Рок КО, Уайт С.З., Джаковски С., Парк Х.В., Леонарди Р. (август 2006 г.). «Прокариотические пантотенаткиназы типа II и III: одна и та же мономерная складка создает димеры с различными каталитическими свойствами». Структура. 14 (8): 1251–61. Дои:10.1016 / j.str.2006.06.008. PMID 16905099.
- ^ а б Бренд LA, Strauss E (май 2005 г.). «Характеристика новой изоформы пантотенаткиназы из Helicobacter pylori». Журнал биологической химии. 280 (21): 20185–8. Дои:10.1074 / jbc.C500044200. PMID 15795230.
- ^ а б Чоудхри А.Е., Мандичак Т.Л., Броски Дж. П., Эгольф Р. В., Кинсленд С., Бегли Т. П., Зеефельд М. А., Ку Т.В., Браун Дж. Р., Залакаин М., Ратнам К. (июнь 2003 г.). «Ингибиторы пантотенаткиназы: новые антибиотики от стафилококковых инфекций». Противомикробные препараты и химиотерапия. 47 (6): 2051–5. Дои:10.1128 / AAC.47.6.2051-2055.2003. ЧВК 155856. PMID 12760898.
- ^ а б Песня WJ, Jackowski S (октябрь 1994 г.). «Кинетика и регуляция пантотенаткиназы из Escherichia coli». Журнал биологической химии. 269 (43): 27051–8. PMID 7929447.
- ^ а б Песня WJ, Jackowski S (октябрь 1992 г.). «Клонирование, секвенирование и экспрессия гена пантотенаткиназы (coaA) Escherichia coli». Журнал бактериологии. 174 (20): 6411–7. Дои:10.1128 / jb.174.20.6411-6417.1992. ЧВК 207592. PMID 1328157.
- ^ а б c Юн М., Парк К.Г., Ким Дж.Й., Rock CO, Джаковски С., Парк Х.В. (сентябрь 2000 г.). «Структурная основа обратной связи регуляции пантотенаткиназы Escherichia coli с помощью кофермента А». Журнал биологической химии. 275 (36): 28093–9. Дои:10.1074 / jbc.M003190200. PMID 10862768.
- ^ Jackowski S, Rock CO (декабрь 1981 г.). «Регуляция биосинтеза кофермента А». Журнал бактериологии. 148 (3): 926–32. ЧВК 216294. PMID 6796563.
- ^ Rock CO, Park HW, Jackowski S (июнь 2003 г.). «Роль регуляции пантотенаткиназы (CoaA) с обратной связью в контроле уровня кофермента А в Escherichia coli». Журнал бактериологии. 185 (11): 3410–5. Дои:10.1128 / JB.185.11.3410-3415.2003. ЧВК 155388. PMID 12754240.
- ^ "Ген PANK2". Домашний справочник по генетике. 2016-02-22. Получено 2016-02-29.
внешние ссылки
- Пантотенат + киназа в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- ЕС 2.7.1.33