Фарнезил-дифосфат фарнезилтрансфераза - Farnesyl-diphosphate farnesyltransferase
| Скваленсинтаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 2.5.1.21 | ||||||||
| Количество CAS | 9077-14-9 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
| фарнезил-дифосфат фарнезилтрансфераза 1 | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Символ | FDFT1 | ||||||
| Ген NCBI | 2222 | ||||||
| HGNC | 3629 | ||||||
| OMIM | 184420 | ||||||
| RefSeq | NM_004462 | ||||||
| UniProt | P37268 | ||||||
| Прочие данные | |||||||
| Номер ЕС | 2.5.1.21 | ||||||
| Locus | Chr. 8 p23.1-p22 | ||||||
| |||||||
Скваленсинтаза (SQS) или фарнезил-дифосфат: фарнезил-дифосфат-фарнезилтрансфераза является фермент локализуется на мембране эндоплазматический ретикулум. SQS участвует в изопреноид биосинтетический путь, катализируя двухступенчатую реакцию, в которой две идентичные молекулы фарнезилпирофосфат (FPP) конвертируются в сквален, с потреблением НАДФН.[2] Катализ от SQS - это первый совершенный шаг в стерол синтез, поскольку полученный сквален преобразуется исключительно в различные стерины, такие как холестерин через сложный, многоэтапный путь. SQS принадлежит семейство сквален / фитоинсинтаза белков.
Разнообразие
Скваленсинтаза была охарактеризована у животных, растений и дрожжей.[3] По своей структуре и механике скваленсинтаза очень похожа на фитоенсинтаза (PHS), другой пренилтрансфераза. PHS выполняет ту же роль, что и SQS у растений и бактерий, катализируя синтез фитоен, предшественник каротиноид соединения.[4]
Структура
Скваленсинтаза (SQS) локализована исключительно в мембрана из эндоплазматический ретикулум (ER).[5] SQS крепится к мембране коротким C-терминал мембранный домен.[6] В N-концевой каталитический домен фермента выступает в цитозоль, где растворимый субстраты связаны.[2] Формы SQS у млекопитающих составляют примерно 47кДа и состоят из ~ 416 аминокислоты. В Кристальная структура SQS человека был определен в 2000 году и показал, что белок полностью состоит из α-спирали. Фермент свернут в единый домен, характеризующийся большим центральным каналом. В активные сайты обе из двух полуреакций, катализируемых SQS, расположены внутри этого канала. Один конец канала открыт для цитозоля, а другой конец образует гидрофобный карман.[5] SQS содержит два сохраненных аспартат -богатые последовательности, которые, как считается, непосредственно участвуют в каталитическом механизме.[7] Эти богатые аспартатом мотивы являются одной из нескольких консервативных структурных особенностей ферментов биосинтеза изопреноидов класса I, хотя эти ферменты не имеют общих последовательностей гомология.[5]

Механизм
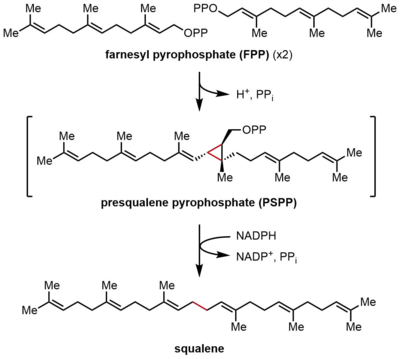
Скваленсинтаза (SQS) катализирует восстановительную димеризацию фарнезилпирофосфата (FPP), при которой две идентичные молекулы FPP превращаются в одну молекулу сквалена. Реакция протекает в две стадии, протекая через промежуточный прескваленпирофосфат (PSPP). FPP - это растворимый аллильный соединение, содержащее 15 атомов углерода (C15), тогда как сквален нерастворим, C30 изопреноид.[2][4] Эта реакция идет лицом к лицу терпен синтез, потому что две молекулы FPP обе соединены в положении C4 и образуют связь 1-1 '. Это контрастирует с 1'-4-связями, которые гораздо более распространены в биосинтезе изопрена, чем 4-4'-связи.[8][9] Механизм реакции SQS требует двухвалентный катион, часто Mg2+, чтобы облегчить привязку пирофосфат группы по FPP.[10]
Конденсация FPP
В первой полуреакции две идентичные молекулы фарнезилпирофосфата (FPP) последовательно связываются со скваленсинтазой (SQS). Молекулы FPP связываются с отдельными участками фермента и имеют разную аффинность связывания.[11] Начиная с вершины каталитического цикла ниже, реакция начинается с ионизации FPP с образованием аллильного карбокатион. А тирозин Остаток (Tyr-171) играет решающую роль на этом этапе, выступая в качестве донора протонов для облегчения извлечения пирофосфата. Более того, образующийся фенолят-анион может стабилизировать образующийся карбокатион за счет катион-π взаимодействия, который был бы особенно сильным из-за богатой электронами природы фенолят-аниона. Образовавшийся аллильный катион затем подвергается атаке олефина второй молекулы FPP, давая третичный карбокатион. Генерированный ранее фенолят-анион затем служит в качестве основы для отделения протона от этого аддукта с образованием циклопропанового продукта, прескваленпирофосфата (PSPP). Созданный PSPP остается связанным с SQS для второй реакции.[5][10] Важность остатка тирозина в этом процессе была продемонстрирована исследованиями мутагенеза с использованием SQS крыс (rSQS),[7] и тем, что Tyr-171 сохраняется во всех известных SQS (и PHS ).[2] В rSQS Tyr-171 был преобразован в ароматические остатки. Phe и Trp, а также гидроксилсодержащий остаток Сер. Ни один из этих мутантов не смог преобразовать FPP в PSPP или сквален, демонстрируя, что одних ароматических колец или спиртов недостаточно для преобразования FPP в PSPP.
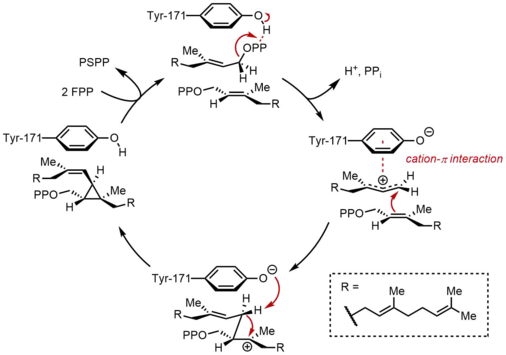
Перестройка и сокращение ПСПП
Во второй полуреакции SQS прескваленпирофосфат (PSPP) перемещается во второй реакционный центр в SQS. Считается, что хранение PSPP в центральном канале SQS защищает реактивный промежуточный продукт от реакции с водой.[5] Из PSPP сквален образуется в результате ряда карбокатионных перегруппировок.[12][13] Процесс начинается с ионизации пирофосфата с образованием циклопропилкарбинильного катиона. Катион перестраивается 1,2-миграция Циклопропановой связи C – C с карбокатионом, образуя связь, показанную синим цветом, с образованием циклобутилкарбокатиона. Впоследствии происходит вторая 1,2-миграция с образованием другого циклопропилкарбинильного катиона, при этом катион находится на третичном атоме углерода. Этот образующийся карбокатион затем открывается гидридом, доставляемым НАДФН, давая сквален, который затем высвобождается SQS в мембрану эндоплазматический ретикулум.[2]
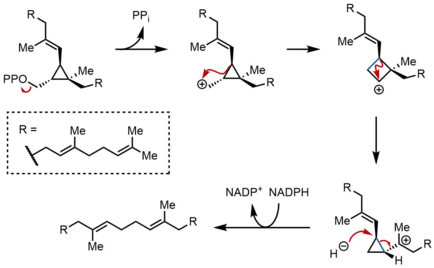
В то время как циклопропилкарбинил-циклопропилкарбинильные перегруппировки могут происходить через дискретные промежуточные циклобутильные катионы, предполагаемый циклобутильный катион не может быть захвачен в модельных исследованиях. Таким образом, катион циклобутила на самом деле может быть переходное состояние между двумя циклопропилкарбинильными катионами, а не дискретным промежуточным соединением. Стереохимия промежуточных продуктов и геометрия олефинов в конечном продукте диктуется надфасциальный характер 1,2-сменной и стереоэлектронные требования. Хотя были предложены другие механизмы, механизм, показанный выше, подтверждается выделением риллингола, который представляет собой спирт, образующийся при захвате второго циклопропилкарбинильного катиона водой.
Регулирование

ГЛП является важным промежуточным звеном в метаболизме мевалонатный путь который представляет собой главную точку ветвления в терпеноид пути.[2][14] FPP используется для образования нескольких важных классов соединений в дополнение к стеролам (через сквален), в том числе убихинон[15] и долихолы.[16] SQS катализирует первый обязательный шаг в биосинтезе стеролов из FPP и поэтому важен для контроля потока в направлении стерол по сравнению с нестериновыми продуктами. Деятельность SQS тесно связана с деятельностью ГМГ-КоА редуктаза, который катализирует лимитирующую стадию мевалонатного пути. Высокий уровень ЛПНП -производный холестерин Значительно подавляют активность HMG-CoA редуктазы, поскольку мевалонат больше не нужен для производства стеролов. Однако остаточная активность HMG-CoA-редуктазы наблюдается даже при очень высоких уровнях LDL, так что FPP может быть использован для образования нестериновых продуктов, необходимых для роста клеток.[17] Чтобы предотвратить использование этого остаточного FPP для синтеза стеролов при большом количестве стеролов, активность SQS значительно снижается, когда уровни LDL высоки.[18] Такое подавление активности SQS лучше рассматривать как механизм контроля потока, а не как способ регулирования уровня холестерина. Это связано с тем, что HMG-CoA-редуктаза является более важным контролирующим фактором для регулирования синтеза холестерина (ее активность подавляется на 98% при высоких уровнях ЛПНП).[17]
Регулирование стеринами
Регулирование SQS происходит в первую очередь на уровне SQS. ген транскрипция.[2] В белок, связывающий регуляторный элемент стерола (SREBP) класс факторы транскрипции играет центральную роль в регуляции генов, участвующих в холестерине гомеостаз, и важен для контроля уровней транскрипции SQS. Когда уровни стеролов низкие, неактивная форма SREBP расщепляется с образованием активного фактора транскрипции, который перемещается в ядро, чтобы вызвать транскрипцию гена SQS. Из трех известных факторов транскрипции SREBP только SREBP-1a и SREBP-2 активируют транскрипцию гена SQS в печени трансгенных мышей.[19][20] В культурном HepG2 клеток, SREBP-1a оказывается более важным, чем SREBP-2, в управлении активацией SQS промоутер.[21] Однако было показано, что промоторы SQS по-разному реагируют на SREBP-1a и SREBP-2 в разных экспериментальных системах.
Помимо SREBP, для максимальной активации промотора SQS необходимы дополнительные факторы транскрипции. Исследования промоутера с использованием люцифераза репортерный ген анализы показал, что Sp1, и NF-Y и / или CREB Факторы транскрипции также важны для активации промотора SQS. NF-Y и / или CREB необходимы для SREBP-1a, чтобы полностью активировать промотор SQS, хотя Sp1 также необходим для SREBP-2 для этого.
Интерактивная карта проезда
Нажмите на гены, белки и метаболиты ниже, чтобы ссылки на соответствующие статьи. [§ 1]

- ^ Интерактивную карту путей можно редактировать на WikiPathways: "Statin_Pathway_WP430".
Биологическая функция
Скваленсинтаза (SQS) - это фермент, участвующий в пути биосинтеза изопреноидов. SQS-синтаза катализирует точку разветвления между биосинтезом стерола и нестерола и связывает фарнезилпирофосфат (FPP) исключительно с образованием стеринов.[2] Важным стеролом, продуцируемым этим путем, является холестерин, который используется в клеточных мембранах и для синтеза гормоны.[22] SQS конкурирует с несколькими другими ферментами за использование FPP, так как он является предшественником множества терпеноидов. Снижение активности SQS ограничивает приток FPP к пути стеролов и увеличивает продукцию нестериновых продуктов. Важные продукты нестерола включают: убихинон, долихолы, гем А, и фарнезилированные белки [23]
Разработка скваленсинтазы нокаутные мыши продемонстрировал, что потеря скваленсинтазы смертельна, и что фермент важен для развития Центральная нервная система.[24]
Актуальность болезни
Скваленсинтаза является мишенью для регуляции уровня холестерина. Вырос выражение SQS повышает уровень холестерина у мышей.[24] Следовательно, ингибиторы SQS представляют большой интерес для лечения гиперхолестеринемия и предотвращение ишемическая болезнь сердца (ИБС).[25] Также было высказано предположение, что варианты этого фермента могут быть частью генетической ассоциации с гиперхолестеринемией.[26]
Ингибиторы скваленсинтазы
Было показано, что ингибиторы скваленсинтазы снижают синтез холестерина, а также уменьшают плазменную триглицерид уровни.[22][27] Ингибиторы SQS могут быть альтернативой Ингибиторы HMG-CoA редуктазы (статины), которые вызывают у некоторых пациентов проблемные побочные эффекты.[28] Ингибиторы скваленсинтазы, которые были исследованы для использования в профилактике сердечно-сосудистые заболевания включают лапаквистат (ТАК-475), сарагозовая кислота и RPR 107393.[29][30] Несмотря на достижение фазы II клинические испытания, производство лапаквистата было прекращено к 2008 году.[31][32]
Ингибирование гомолога скваленсинтазы в Золотистый стафилококк в настоящее время исследуется как фактор вирулентности антибактериальный терапия.[33]
Модельные организмы
Модельные организмы были использованы при изучении функции FDFT1. Условный нокаутирующая мышь линия называется Fdft1tm1a (КОМП) Wtsi был создан на Wellcome Trust Sanger Institute.[34] Самцы и самки животных прошли стандартизованный фенотипический скрининг[35] для определения последствий удаления.[36][37][38][39] Проведены дополнительные проверки: - Углубленное иммунологическое фенотипирование[40]
| Характеристика | Фенотип |
|---|---|
| Все данные доступны на сайте.[35][40] | |
| Гематология 6 недель | Нормальный |
| Инсулин | Нормальный |
| Гомозиготная жизнеспособность на P14 | Аномальный |
| Рецессивный смертельное исследование | Аномальный |
| Масса тела | Нормальный |
| Неврологический осмотр | Нормальный |
| Сила захвата | Нормальный |
| Дисморфология | Нормальный |
| Косвенная калориметрия | Нормальный |
| Тест толерантности к глюкозе | Нормальный |
| Слуховой ответ ствола мозга | Нормальный |
| DEXA | Нормальный |
| Рентгенография | Нормальный |
| Морфология глаза | Нормальный |
| Клиническая химия | Нормальный |
| Гематология 16 недель | Нормальный |
| Лейкоциты периферической крови 16 недель | Нормальный |
| Вес сердца | Нормальный |
| Сальмонелла инфекция | Нормальный |
| Функция цитотоксических Т-клеток | Нормальный |
| Иммунофенотипирование селезенки | Нормальный |
| Иммунофенотипирование мезентериальных лимфатических узлов | Нормальный |
| Иммунофенотипирование костного мозга | Нормальный |
| Состав эпидермального иммунитета | Нормальный |
| Тричурис Вызов | Нормальный |
использованная литература
- ^ Итикава М., Йокомидзо А., Ито М., Сугита К., Усуи Х., Симидзу Х., Судзуки М., Тераяма К., Канда А. (март 2011 г.). «Открытие новой матрицы 2-аминобензгидрола для сильнодействующих ингибиторов скваленсинтазы». Биоорг. Med. Chem. 19 (6): 1930–49. Дои:10.1016 / j.bmc.2011.01.065. PMID 21353782.
- ^ а б c d е ж г час Tansey TR, Shechter I (декабрь 2000 г.). «Структура и регуляция скваленсинтазы млекопитающих». Биохим. Биофиз. Acta. 1529 (1–3): 49–62. Дои:10.1016 / S1388-1981 (00) 00137-2. PMID 11111077.
- ^ Накашима Т., Иноуэ Т., Ока А., Нишино Т., Осуми Т., Хата С. (март 1995 г.). «Клонирование, экспрессия и характеристика кДНК, кодирующих скваленсинтазу Arabidopsis thaliana». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 92 (6): 2328–32. Bibcode:1995ПНАС ... 92.2328Н. Дои:10.1073 / пнас.92.6.2328. ЧВК 42476. PMID 7892265.
- ^ а б Танси Т.Р., Шехтер I (2001). Скваленсинтаза: структура и регуляция. Прог. Nucleic Acid Res. Мол. Биол. Прогресс в исследованиях нуклеиновых кислот и молекулярной биологии. 65. С. 157–95. Дои:10.1016 / S0079-6603 (00) 65005-5. ISBN 9780125400657. PMID 11008488.
- ^ а б c d е Пандит Дж., Дэнли Д.Е., Шульте Г.К., Маззалупо С., Поли Т.А., Хейворд С.М., Хаманака Е.С., Томпсон Дж.Ф., Харвуд Г.Дж. (сентябрь 2000 г.). «Кристаллическая структура скваленсинтазы человека. Ключевой фермент биосинтеза холестерина». J. Biol. Chem. 275 (39): 30610–7. Дои:10.1074 / jbc.M004132200. PMID 10896663.
- ^ Дженнингс С.М., Цай Ю.Х., Фиш Т.М., Робинсон Г.В. (июль 1991 г.). «Молекулярное клонирование и характеристика дрожжевого гена скваленсинтетазы». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 88 (14): 6038–42. Bibcode:1991PNAS ... 88.6038J. Дои:10.1073 / pnas.88.14.6038. ЧВК 52017. PMID 2068081.
- ^ а б Гу П, Исии Й, Спенсер Т.А., Шехтер I. (май 1998 г.). «Исследование функциональной структуры и идентификация трех ферментных доменов, участвующих в каталитической активности скваленсинтазы печени крыс». J. Biol. Chem. 273 (20): 12515–25. Дои:10.1074 / jbc.273.20.12515. PMID 9575210.
- ^ Поултер CD (1990). «Биосинтез терпенов не голова к хвосту. Формирование 1'-1 и 1'-3 связей». Отчеты о химических исследованиях. 23 (3): 70–77. Дои:10.1021 / ar00171a003.
- ^ Линь Ф.Й., Лю К.И., Лю Ю.Л., Чжан Ю., Ван К., Дженг В.Й., Ко Т.П., Цао Р., Ван А.Х., Олдфилд Е. (декабрь 2010 г.). «Механизм действия и ингибирование дегидроскваленсинтазы». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 107 (50): 21337–42. Bibcode:2010PNAS..10721337L. Дои:10.1073 / pnas.1010907107. ЧВК 3003041. PMID 21098670.
- ^ а б Бейтия Э., Куреши А.А., Портер Дж.В. (март 1973 г.). «Скваленсинтетаза. 3. Механизм реакции». J. Biol. Chem. 248 (5): 1856–67. PMID 4348553.
- ^ Мухтияр К.А., Калиновский С.С., Чжан Д., Поултер CD (апрель 1994 г.). «Скваленсинтаза дрожжей. Механизм добавления субстратов и активации НАДФН». J. Biol. Chem. 269 (15): 11201–7. PMID 8157649.
- ^ Blagg, Брайан С. Дж .; Ярстфер, Майкл Б .; Роджерс, Дэниел Х .; Поултер, К. Дейл (2002-07-04). «Рекомбинантная сквален-синтаза. Механизм перегруппировки пресквалендифосфата в сквален». Журнал Американского химического общества. 124 (30): 8846–8853. Дои:10.1021 / ja020411a. PMID 12137537.
- ^ Ярстфер, Майкл Б .; Blagg, Брайан С. Дж .; Роджерс, Дэниел Х .; Поултер, К. Дейл (1996-12-25). "Биосинтез сквалена. Доказательства третичного циклопропилкарбинилового катионного промежуточного соединения в перегруппировке пресквалендифосфата в сквален". Журнал Американского химического общества. 118 (51): 13089–13090. Дои:10.1021 / ja963308s.
- ^ Браун, Майкл С .; Гольдштейн, Джозеф Л. (1980). «Поливалентная обратная связь регуляции HMG CoA редуктазы, механизма контроля, координирующего синтез изопреноидов и рост клеток». J. Lipid Res. 21 (5): 505–517. PMID 6995544.
- ^ Олсон, Роберт Э. (1967-01-01). Роберт С. Харрис, Ира Г. Вул, Джон А. Лорейн, Дж. Ф. Марриан и Кеннет В. Тиманн (ред.). «Биосинтез убихинонов у животных *». Витамины и гормоны. 24: 551–574. Дои:10.1016 / s0083-6729 (08) 60221-6. ISBN 9780127098241. PMID 5340877.
- ^ Gough, Doreen P .; Хемминг, Ф. В. (1970-06-01). «Характеристика и стереохимия биосинтеза долихолов в печени крыс». Биохимический журнал. 118 (1): 163–166. Дои:10.1042 / bj1180163. ISSN 0264-6021. ЧВК 1179092. PMID 4319540.
- ^ а б Фауст, Джерри Р .; Goldstein, Joseph L .; Браун, Майкл С. (1979-01-01). «Синтез убихинона и холестерина в человеческих фибробластах: регуляция разветвленного пути». Архивы биохимии и биофизики. 192 (1): 86–99. Дои:10.1016/0003-9861(79)90074-2. PMID 219777.
- ^ Фауст, Джерри Р .; Goldstein, Joseph L .; Браун, Майкл С. (1979-10-01). «Активность скваленсинтетазы в человеческих фибробластах: регулирование через рецептор липопротеинов низкой плотности». Труды Национальной академии наук Соединенных Штатов Америки. 76 (10): 5018–5022. Bibcode:1979PNAS ... 76.5018F. Дои:10.1073 / pnas.76.10.5018. ISSN 0027-8424. ЧВК 413070. PMID 228272.
- ^ Guan, G .; Jiang, G .; Koch, R. L .; Шехтер, И. (1995-09-15). «Молекулярное клонирование и функциональный анализ промотора гена скваленсинтазы человека». Журнал биологической химии. 270 (37): 21958–21965. Дои:10.1074 / jbc.270.37.21958. ISSN 0021-9258. PMID 7665618.
- ^ Гуань, Гуйминь; Дай, Пей-Хуа; Осборн, Тимоти Ф .; Kim, Jae B .; Шехтер, Исайаху (11 апреля 1997 г.). «Множественные элементы последовательности вовлечены в регуляцию транскрипции гена скваленсинтазы человека». Журнал биологической химии. 272 (15): 10295–10302. Дои:10.1074 / jbc.272.15.10295. ISSN 0021-9258. PMID 9092581.
- ^ Guan, G .; Dai, P .; Шехтер, И. (1998-05-15). «Дифференциальная регуляция транскрипции гена скваленсинтазы человека с помощью белков, связывающих регуляторный элемент стерола (SREBP) 1a и 2, и участие 5'-элементов последовательности ДНК в регуляции». Журнал биологической химии. 273 (20): 12526–12535. Дои:10.1074 / jbc.273.20.12526. ISSN 0021-9258. PMID 9575211.
- ^ а б Куруунакис А.П., Кацелу М.Г., Матралис А.Н., Ладопулу Е.М., Бававеа Э. (2011). «Ингибиторы скваленсинтазы: обновленная информация о поиске новых антигиперлипидемических и антиатеросклеротических агентов». Curr. Med. Chem. 18 (29): 4418–39. Дои:10.2174/092986711797287557. PMID 21864285.
- ^ Парадайз ЭМ, Кирби Дж., Чан Р., Кислинг Дж. Д. (июнь 2008 г.). «Перенаправление потока через точку ветвления FPP в Saccharomyces cerevisiae путем подавления скваленсинтазы». Biotechnol. Bioeng. 100 (2): 371–8. Дои:10.1002 / бит. 21766. PMID 18175359. S2CID 23878922.
- ^ а б Окадзаки Х., Тадзо Ф., Окадзаки С., Ису Н., Цукамото К., Секия М., Яхаги Н., Иидзука Й, Охаши К., Китамине Т, Тодзава Р., Инаба Т, Ягю Х, Окадзаки М., Шимано Х, Сибата Н., Араи Х. , Нагаи Р.З., Кадоваки Т., Осуга Дж., Ишибаши С. (сентябрь 2006 г.). «Повышенный биосинтез холестерина и гиперхолестеринемия у мышей со сверхэкспрессией скваленсинтазы в печени». J. Lipid Res. 47 (9): 1950–8. Дои:10.1194 / мл. M600224-JLR200. PMID 16741291.
- ^ Дэвидсон MH (январь 2007 г.). «Ингибирование скваленсинтазы: новая цель для лечения дислипидемии». Карр Атеросклер Представитель. 9 (1): 78–80. Дои:10.1007 / BF02693932. PMID 17169251. S2CID 28176904.
- ^ Do R, Kiss RS, Gaudet D, Engert JC (январь 2009 г.). «Скваленсинтаза: критический фермент в пути биосинтеза холестерина». Clin. Genet. 75 (1): 19–29. Дои:10.1111 / j.1399-0004.2008.01099.x. PMID 19054015. S2CID 205406994.
- ^ Хиёси Х., Янагимачи М., Ито М., Саеки Т., Йошида И., Окада Т., Икута Х., Синмё Д., Танака К., Курусу Н., Танака Х. (ноябрь 2001 г.). «Ингибиторы скваленсинтазы снижают уровень триглицеридов плазмы посредством независимого от рецепторов липопротеинов низкой плотности механизма». Евро. J. Pharmacol. 431 (3): 345–52. Дои:10.1016 / S0014-2999 (01) 01450-9. PMID 11730728.
- ^ Сейки С., Фришман WH (2009). «Фармакологическое ингибирование скваленсинтазы и других нижестоящих ферментов пути синтеза холестерина: новый терапевтический подход к лечению гиперхолестеринемии». Cardiol Rev. 17 (2): 70–6. Дои:10.1097 / CRD.0b013e3181885905. PMID 19367148. S2CID 33130333.
- ^ Чарльтон-Менис V, Даррингтон П.Н. (2007). «Ингибиторы скваленсинтазы: клиническая фармакология и потенциал снижения уровня холестерина». Наркотики. 67 (1): 11–6. Дои:10.2165/00003495-200767010-00002. PMID 17209661. S2CID 45715717.
- ^ Амин Д., Ратледж Р.З., Нидл С.Н., Гальценски Х.Ф., Нойеншвандер К., Скотез А.С., Магуайр М.П., Буш Р.С., Хеле Д.Д., Билдер Г.Е., Перрон М.Х. (май 1997 г.). «RPR 107393, мощный ингибитор скваленсинтазы и эффективный при пероральном приеме холестерин-снижающий агент: сравнение с ингибиторами HMG-CoA редуктазы». J. Pharmacol. Exp. Ther. 281 (2): 746–52. PMID 9152381.
- ^ Гиббс, Эдвина (29 октября 2007 г.). «ОБНОВЛЕНИЕ 2 - FDA США говорит Takeda прекратить некоторые испытания TAK-475». Рейтер. Получено 5 марта 2013.
- ^ «Прекращение разработки TAK-475, соединения для лечения гиперхолестеринемии». Такеда Фармацевтическая Компания Лимитед. 28 марта 2008 г.. Получено 5 марта 2013.
- ^ Лю CI, Лю GY, Song Y, Yin F, Hensler ME, Jeng WY, Nizet V, Wang AH, Oldfield E (март 2008 г.). «Ингибитор биосинтеза холестерина блокирует вирулентность Staphylococcus aureus» (PDF). Наука. 319 (5868): 1391–4. Bibcode:2008Научный ... 319.1391L. Дои:10.1126 / science.1153018. ЧВК 2747771. PMID 18276850.
- ^ Гердин А.К. (2010). «Программа генетики Sanger Mouse: характеристика мышей с высокой пропускной способностью». Acta Ophthalmologica. 88: 925–7. Дои:10.1111 / j.1755-3768.2010.4142.x. S2CID 85911512.
- ^ а б «Международный консорциум по фенотипированию мышей».
- ^ Скарнес В.К., Розен Б., Вест А.П., Кутсуракис М., Бушелл В., Айер В., Мухика А.О., Томас М., Харроу Дж., Кокс Т., Джексон Д., Северин Дж., Биггс П., Фу Дж., Нефедов М., де Йонг П.Дж., Стюарт AF, Брэдли А. (июнь 2011 г.). «Ресурс условного нокаута для полногеномного исследования функции генов мыши». Природа. 474 (7351): 337–42. Дои:10.1038 / природа10163. ЧВК 3572410. PMID 21677750.
- ^ Долгин Э (июнь 2011 г.). "Библиотека мыши настроена на нокаут". Природа. 474 (7351): 262–3. Дои:10.1038 / 474262a. PMID 21677718.
- ^ Коллинз Ф.С., Россант Дж., Вурст В. (январь 2007 г.). «Мышь по всем причинам». Ячейка. 128 (1): 9–13. Дои:10.1016 / j.cell.2006.12.018. PMID 17218247. S2CID 18872015.
- ^ White JK, Gerdin AK, Karp NA, Ryder E, Buljan M, Bussell JN, Salisbury J, Clare S, Ingham NJ, Podrini C, Houghton R, Estabel J, Bottomley JR, Melvin DG, Sunter D, Adams NC, Sanger Institute Проект генетики мышей, Таннахилл Д., Логан Д.В., Макартур Д.Г., Флинт Дж., Махаджан В.Б., Цанг С.Х., Смит I, Ватт FM, Скарнес В.К., Дуган Джи, Адамс DJ, Рамирес-Солис Р., Брэдли А., Сталь КП (2013) . «Полногеномное поколение и систематическое фенотипирование мышей с нокаутом открывает новые роли для многих генов». Ячейка. 154 (2): 452–64. Дои:10.1016 / j.cell.2013.06.022. ЧВК 3717207. PMID 23870131.
- ^ а б «Консорциум иммунофенотипирования инфекций и иммунитета (3i)».
внешние ссылки
- Фарнезил-дифосфат + фарнезилтрансфераза в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)