Орнитин декарбоксилаза - Ornithine decarboxylase
| орнитиндекарбоксилаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Димер орнитиндекарбоксилазы, человек | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 4.1.1.17 | ||||||||
| Количество CAS | 9024-60-6 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| BRENDA | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
| орнитиндекарбоксилаза | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Символ | ODC1 | ||||||
| Ген NCBI | 4953 | ||||||
| HGNC | 8109 | ||||||
| OMIM | 165640 | ||||||
| RefSeq | NM_002539 | ||||||
| UniProt | P11926 | ||||||
| Прочие данные | |||||||
| Номер ЕС | 4.1.1.17 | ||||||
| Locus | Chr. 2 p25 | ||||||
| |||||||
Фермент орнитиндекарбоксилаза (ODC) катализирует декарбоксилирование из орнитин (продукт цикл мочевины ) сформировать путресцин. Эта реакция - совершенный шаг в полиамин синтез.[1] У человека этот белок насчитывает 461 аминокислоты и образует гомодимер.
Механизм реакции
Лизин 69 на орнитиндекарбоксилазе (ODC) связывает кофактор пиридоксальфосфат сформировать База Шиффа. Орнитин замещает лизин с образованием основания Шиффа, присоединенного к ортонину, которое декарбоксилатируется с образованием хиноид промежуточный. Это промежуточное звено перестраивается, чтобы сформировать базу Шиффа, прикрепленную к путресцин, на которую нападают лизин для выпуска продукта путресцина и преобразования ODC, связанного с PLP.[2]

Это первый шаг и шаг, ограничивающий скорость производства у людей полиамины, соединения, необходимые для деление клеток.
Структура
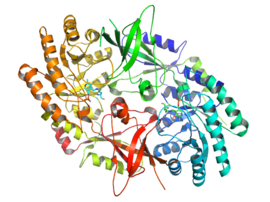
Активная форма орнитиндекарбоксилазы - это гомодимер. Каждый мономер содержит бочкообразный домен, состоящий из альфа-бета ствол, и листовой домен, состоящий из двух бета-листы. Домены соединены петлями. Мономеры соединяются друг с другом посредством взаимодействия между цилиндром одного мономера и листом другого. Связывание между мономерами относительно слабое, и ODC быстро преобразуется между мономерными и димерными формами в клетке.[1]
В пиридоксальфосфат кофактор связывает лизин 69 на C-конец конец домена ствола. В активный сайт находится на границе раздела двух доменов в полости, образованной петлями из обоих мономеров.[1]
Функция
Реакция декарбоксилирования орнитина, катализируемая орнитиндекарбоксилазой, является первой и совершенный шаг в синтезе полиамины особенно путресцин, спермидин и спермин. Полиамины важны для стабилизации ДНК структура, ДНК ремонт с двойным разрывом цепи путь и как антиоксиданты. Следовательно, орнитиндекарбоксилаза является важным ферментом для роста клеток, производящим полиамины, необходимые для стабилизации вновь синтезированной ДНК. Отсутствие ODC вызывает клеточную апоптоз у эмбриональных мышей, вызванное повреждением ДНК.[4]
Протеасомная деградация
ODC - это наиболее хорошо охарактеризованный клеточный белок, подверженный убиквитин -независимый протеасомный деградация. Хотя большинство белков сначала необходимо пометить несколькими убиквитин молекулы, прежде чем они будут связаны и разложены протеасома, Деградация ODC вместо этого опосредуется несколькими сайтами узнавания на белке и его вспомогательном факторе. антизим. Процесс деградации ODC регулируется в петля отрицательной обратной связи продуктами его реакции.[5]
До отчета Sheaff et al. (2000),[6] которые продемонстрировали, что ингибитор циклин-зависимой киназы (Cdk) p21Cip1 также деградируется протеасомой убиквитин-независимым образом, ODC был единственным ярким примером убиквитин-независимой протеасомной деградации.[7]
Клиническое значение
ODC является транскрипционной мишенью онкоген Мой с[8] и активируется при большом количестве видов рака. Полиаминовые продукты пути, инициализированного ODC, связаны с усилением роста клеток и снижением апоптоз.[9] Ультрафиолетовое излучение,[10] асбест[11] и андрогены выпущен простата[12] известно, что они вызывают повышенную активность ODC, связанную с раком. Ингибиторы ODC, такие как эфлорнитин было показано, что эффективно снижает риск рака на животных моделях,[13] и препараты, нацеленные на ODC, проходят испытания для потенциального клинического использования. Механизм, с помощью которого ODC способствует канцерогенез сложна и не совсем известна. Помимо прямого влияния на стабильность ДНК, полиамины также активируют щелевой переход гены[14] и подавлять плотные контакты гены. Гены щелевых соединений участвуют в коммуникации между канцерогенными клетками, а гены плотных соединений действуют как супрессоры опухолей.[9]
Экспрессия гена ODC индуцируется большим количеством биологических стимулов, включая захват активность в головном мозге.[15] Инактивация ODC дифторметилорнитином (эфлорнитин ) используется для лечения рак и рост волос на лице у женщин в постменопаузе.
ODC также является незаменимым ферментом для паразиты любить трипаносома, лямблии, и плазмодий, факт, эксплуатируемый наркотиками эфлорнитин.[16]
использованная литература
- ^ а б c Керн А.Д., Оливейра М.А., Коффино П., Хакерт М.Л. (май 1999 г.). «Структура орнитиндекарбоксилазы млекопитающих при разрешении 1,6 A: стереохимические последствия PLP-зависимых декарбоксилаз аминокислот». Структура. 7 (5): 567–81. Дои:10.1016 / S0969-2126 (99) 80073-2. PMID 10378276.
- ^ Brooks HB, Phillips MA (декабрь 1997 г.). «Характеристика механизма реакции орнитиндекарбоксилазы Trypanosoma brucei с помощью многоволновой спектроскопии остановленного потока». Биохимия. 36 (49): 15147–55. Дои:10.1021 / bi971652b. PMID 9398243.
- ^ PDB: 1d7k; Альмруд Дж. Дж., Оливейра М. А., Керн А. Д., Гришин Н. В., Филлипс М. А., Хакерт М. Л. (январь 2000 г.). «Кристаллическая структура орнитиндекарбоксилазы человека при разрешении 2,1 A: структурное понимание связывания антизима». J. Mol. Биол. 295 (1): 7–16. Дои:10.1006 / jmbi.1999.3331. PMID 10623504.; предоставлено через PyMOL.
- ^ Пендевиль Х., Карпино Н., Марин Дж. К. и др. (Октябрь 2001 г.). «Ген орнитиндекарбоксилазы необходим для выживания клеток на раннем этапе развития мышей» (PDF). Мол. Cell. Биол. 21 (19): 6549–58. Дои:10.1128 / MCB.21.19.6549-6558.2001. ЧВК 99801. PMID 11533243.
- ^ Чжан М., Пикарт С.М., Коффино П. (апрель 2003 г.). «Детерминанты распознавания протеасомами орнитиндекарбоксилазы, убиквитиннезависимого субстрата». EMBO J. 22 (7): 1488–96. Дои:10.1093 / emboj / cdg158. ЧВК 152902. PMID 12660156.
- ^ Sheaff RJ, Singer JD, Swanger J, Smitherman M, Roberts JM, Clurman BE (февраль 2000 г.). «Протеасомный оборот p21Cip1 не требует убиквитинирования p21Cip1». Мол. Ячейка. 5 (2): 403–10. Дои:10.1016 / S1097-2765 (00) 80435-9. PMID 10882081.
- ^ Verma R, Deshaies RJ (май 2000 г.). «Протеасомный Howdunit: случай отсутствия сигнала». Ячейка. 101 (4): 341–4. Дои:10.1016 / S0092-8674 (00) 80843-0. PMID 10830160. S2CID 18425370.
- ^ Белло-Фернандес С., Пакхэм Дж., Кливленд Дж. Л. (август 1993 г.). «Ген орнитиндекарбоксилазы является транскрипционной мишенью c-Myc». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 90 (16): 7804–8. Дои:10.1073 / пнас.90.16.7804. ЧВК 47231. PMID 8356088.
- ^ а б Гернер Е.В., Мейскенс Флорида (октябрь 2004 г.). «Полиамины и рак: старые молекулы, новое понимание». Nat. Преподобный Рак. 4 (10): 781–92. Дои:10.1038 / nrc1454. PMID 15510159. S2CID 37647479.
- ^ Ахмад Н., Гиллиам А.С., Катияр С.К., О'Брайен Т.Г., Мухтар Х. (сентябрь 2001 г.). «Решающая роль орнитиндекарбоксилазы в фотоканцерогенезе». Am. Дж. Патол. 159 (3): 885–92. Дои:10.1016 / S0002-9440 (10) 61764-6. ЧВК 1850478. PMID 11549581.
- ^ Марш Дж. П., Моссман Б. Т. (январь 1991 г.). «Роль асбеста и активных форм кислорода в активации и экспрессии орнитиндекарбоксилазы в эпителиальных клетках трахеи хомяка». Рак Res. 51 (1): 167–73. PMID 1846307.
- ^ Crozat A, Palvimo JJ, Julkunen M, Jänne OA (март 1992 г.). «Сравнение андрогенной регуляции экспрессии генов орнитиндекарбоксилазы и S-аденозилметиониндекарбоксилазы в почках грызунов и дополнительных половых органах». Эндокринология. 130 (3): 1131–44. Дои:10.1210 / en.130.3.1131. PMID 1537280.
- ^ Мейскенс Флорида, Гернер Э.В. (май 1999 г.). «Разработка дифторметилорнитина (DFMO) в качестве химиопрофилактического агента». Clin. Рак Res. 5 (5): 945–51. PMID 10353725.
- ^ Шор Л., Маклин П., Гилмор С.К., Ходгинс МБ, Финбоу М.Э. (июль 2001 г.). «Полиамины регулируют связь щелевых соединений в клетках, экспрессирующих коннексин 43». Biochem. J. 357 (Pt 2): 489–95. Дои:10.1042/0264-6021:3570489. ЧВК 1221976. PMID 11439099.
- ^ Герберг LJ, Rose IC, de Belleroche JS, Mintz M (1992). «Индукция орнитиндекарбоксилазы и синтез полиамина при киндлинге припадков: эффект альфа-дифторметилорнитина». Эпилепсия Res. 11 (1): 3–7. Дои:10.1016 / 0920-1211 (92) 90015-Л. PMID 1563337. S2CID 1221264.
- ^ Heby O, Persson L, Rentala M (август 2007 г.). «Ориентация на ферменты биосинтеза полиамина: многообещающий подход к терапии африканской сонной болезни, болезни Шагаса и лейшманиоза». Аминокислоты. 33 (2): 359–66. Дои:10.1007 / s00726-007-0537-9. PMID 17610127. S2CID 26273053.
внешние ссылки
- Орнитин декарбоксилаза на herkules.oulu.fi
- Орнитин + декарбоксилаза в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)