Эндонуклеаза - Endonuclease
Эндонуклеазы находятся ферменты которые раскалывают фосфодиэфирная связь в пределах полинуклеотид цепь. Некоторые, например дезоксирибонуклеаза I, разрезают ДНК относительно неспецифично (безотносительно к последовательности), в то время как многие, как правило, эндонуклеазы рестрикции или рестрикционные ферменты, расщепляют только очень специфические нуклеотидные последовательности. Эндонуклеазы отличаются от экзонуклеазы, которые расщепляют концы узнаваемых последовательностей вместо средней (эндо) части. Некоторые ферменты, известные как "экзоэндонуклеазы", однако, не ограничиваются какой-либо функцией нуклеазы, проявляя качества, которые являются как эндо-, так и экзо-подобными.[1] Данные свидетельствуют о том, что активность эндонуклеазы отстает от активности экзонуклеазы.[2]
Ферменты рестрикции эндонуклеазы из эубактерии и археи которые распознают определенную последовательность ДНК.[3] Нуклеотидная последовательность, распознаваемая для расщепления рестрикционным ферментом, называется сайтом рестрикции. Обычно сайт рестрикции представляет собой палиндромный последовательность длиной от четырех до шести нуклеотидов. Большинство эндонуклеаз рестрикции расщепляют цепь ДНК неравномерно, оставляя комплементарные одноцепочечные концы. Эти концы могут повторно соединяться посредством гибридизации и называются «липкими концами». После образования пары фосфодиэфирные связи фрагментов могут быть соединены ДНК-лигаза. Известны сотни эндонуклеаз рестрикции, каждая из которых атакует свой сайт рестрикции. Фрагменты ДНК, расщепленные одной и той же эндонуклеазой, могут быть соединены вместе независимо от происхождения ДНК. Такая ДНК называется рекомбинантная ДНК; ДНК образуется в результате объединения генов в новые комбинации.[4] Эндонуклеазы рестрикции (рестрикционные ферменты ) делятся на три категории: Тип I, Тип II и Тип III, в соответствии с их механизмом действия. Эти ферменты часто используются в генная инженерия сделать рекомбинантная ДНК для введения в клетки бактерий, растений или животных, а также в синтетическая биология.[5] Одна из самых известных эндонуклеаз - Cas9.
Категории
В конечном счете, есть три категории эндонуклеазы рестрикции которые относительно способствуют расщеплению определенных последовательностей. Типы I и III представляют собой большие мультисубъединичные комплексы, которые включают как эндонуклеазы и метилаза виды деятельности. Тип I может отщеплять на случайных участках около 1000 пар оснований или более из последовательности распознавания, и для этого требуется АТФ в качестве источника энергии. Тип II ведет себя несколько иначе и был впервые выделен Гамильтоном Смитом в 1970 году. Это более простые версии эндонуклеаз, которым не требуется АТФ в процессах разложения. Некоторые примеры эндонуклеаз рестрикции типа II включают: БамЗДРАВСТВУЙ, ЭкоRI, ЭкоRV, HindIII, и ХэIII. Тип III, однако, расщепляет ДНК примерно на 25 пар оснований из последовательности распознавания и также требует АТФ в этом процессе.[4]
Обозначения
Обычно используемые обозначения эндонуклеаз рестрикции[6] имеет форму "VwxyZ ", где"Vwx«курсивом выделены первая буква названия рода и первые две буквы вида, в котором может быть обнаружена эта эндонуклеаза рестрикции, например, кишечная палочка, Эко, и Haemophilus influenzae, Hin. За ним следует необязательный символ «y» без курсива, который указывает тип или идентификацию штамма, например, ЭкоR для Кишечная палочка штаммы, несущие фактор передачи лекарственной устойчивости RTF-1,[6] ЭкоB для Кишечная палочка штамм B,[7] и Hind для H. influenzae напряжение d.[6] Наконец, когда конкретный тип или штамм имеет несколько различных эндонуклеаз рестрикции, они обозначаются римскими цифрами, таким образом, эндонуклеазы рестрикции из H. influenzae штамм d назван HindI, HindII, HindIII и т. д. Другой пример: "ХэII "и"ХэIII "относятся к бактерии Haemophilus aegyptius (штамм не указан), эндонуклеазы рестрикции номер II и номер III соответственно.[4]:64–64 Рестрикционные ферменты, используемые в молекулярной биологии, обычно распознают короткие последовательности-мишени примерно из 4-8 пар оснований. Например, ЭкоФермент RI распознает и расщепляет последовательность 5 '- GAATTC - 3'.[8]
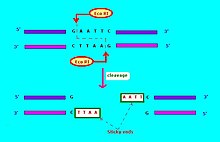
Эндонуклеазы рестрикции бывают нескольких типов. Эндонуклеаза рестрикции обычно требует сайта узнавания и образца расщепления (обычно нуклеотидных оснований: A, C, G, T). Если сайт узнавания находится за пределами области паттерна расщепления, то эндонуклеаза рестрикции называется типом I. Если последовательность узнавания перекрывается с последовательностью расщепления, то эндонуклеаза рестрикции рестрикционный фермент это Тип II.
Дальнейшее обсуждение
Могут быть обнаружены рестрикционные эндонуклеазы, которые расщепляют стандартную дцДНК (двухцепочечную ДНК), или оцДНК (одноцепочечную ДНК), или даже РНК. Это обсуждение ограничивается дцДНК; тем не менее, обсуждение может быть расширено до следующего:
- Стандартная дцДНК
- Нестандартная ДНК
- Праздничные развязки
- Трехцепочечная ДНК, четырехцепочечная ДНК (G-квадруплекс )
- Двухцепочечные гибриды ДНК и РНК (одна цепь - ДНК, другая - РНК)[4]:72–73
- Синтетическая или искусственная ДНК (например, содержащая основания, отличные от A, C, G, T, относится к работе Эрик Т. Кул ). Исследования с синтетическими кодоны, обратитесь к исследованию S. Benner и увеличив аминокислотный набор в полипептидах, таким образом увеличив протеом или протеомика см. исследование П. Шульца.[4]:Глава 3
Кроме того, в настоящее время ведутся исследования по созданию синтетических или искусственных эндонуклеаз рестрикции, особенно с сайтами узнавания, которые уникальны в пределах генома.
Эндонуклеазы рестрикции или рестрикционные ферменты обычно раскалываются двумя способами: узоры с тупыми или липкими концами. Пример эндонуклеазы рестрикции типа I.[4]:64
Кроме того, существуют ДНК / РНК неспецифические эндонуклеазы, такие как те, которые находятся в Serratia marcescens, которые действуют на дцДНК, оцДНК и РНК.
Ремонт ДНК
Эндонуклеазы играют роль в репарации ДНК. Эндонуклеаза AP, в частности, катализирует разрез ДНК исключительно в AP-сайтах и, следовательно, подготавливает ДНК для последующего вырезания, репарационного синтеза и лигирования ДНК. Например, когда происходит депуринизация, это поражение оставляет сахар дезоксирибозы с отсутствующим основанием.[9] Эндонуклеаза АР распознает этот сахар и по существу разрезает ДНК на этом участке, а затем позволяет продолжить восстановление ДНК.[10] Кишечная палочка клетки содержат две AP-эндонуклеазы: эндонуклеазу IV (endoIV) и экзонуклеазу III (exoIII), тогда как у эукариот существует только одна AP-эндонуклеаза.[11]

Общие эндонуклеазы
Ниже приведены таблицы распространенных прокариотических и эукариотических эндонуклеаз.[12]
| Прокариотический фермент | Источник | Комментарии |
|---|---|---|
| RecBCD энонуклеаза | Кишечная палочка | Частично зависит от АТФ; также экзонуклеаза; функции в рекомбинации и ремонте |
| Эндонуклеаза Т7 (P00641) | фаг Т7 (ген 3) | Необходим для репликации; предпочтение одноцепочечной ДНК над двухцепочечной |
| Эндонуклеаза Т4 II (P07059) | фаг Т4 (denA) | Расщепляет последовательность -TpC- с получением олигонуклеотидов с концевым 5'-dCMP; длина цепочки продукта зависит от условий |
| Эндонуклеаза Bal 31 | P. espejiana | Также экзонуклеаза; откусывает 3 'и 5' концы дуплексной ДНК. Смесь как минимум двух нуклеаз, быстрой и медленной.[13] |
| Эндонуклеаза I (эндо I; P25736) | E. coli (endA) | Периплазматическая локация; средняя длина цепочки товара 7; ингибируется тРНК; производит двухцепочечный разрыв ДНК; производит ник при образовании комплекса с тРНК; endo I мутанты нормально растут |
| Микрококковая нуклеаза (P00644) | Стафилококк | Образует 3'-P концы; требуется Ca2 +; также действует на РНК; предпочитает одноцепочечную ДНК и богатые АТ участки |
| Эндонуклеаза II (эндо VI, экзо III; P09030) | E. coli (xthA) | Расщепление рядом с AP-сайтом; также экзонуклеаза 3 '-> 5'; фосфомоноэстераза на 3'-P концах |
| Эукариотический фермент | Источник | Комментарии |
| Эндонуклеаза нейроспоры[14] | Neurospora crassa, митохондрии | Также действует на РНК. |
| Нуклеаза S1 (P24021) | Aspergillus oryzae | Также действует на РНК |
| Р1-нуклеаза (P24289) | Penicillium citrinum | Также действует на РНК |
| Нуклеаза бобов мунг я | ростки маша | Также действует на РНК |
| Нуклеаза устилаго (ДНКаза I)[15] | Ustilago Maydis | Также действует на РНК |
| Дназа I (P00639) | Поджелудочная железа крупного рогатого скота | Средняя длина цепочки товара - 4; образует двойной разрыв цепи в присутствии Mn2 + |
| Эндонуклеаза AP | Ядро, митохондрии | Участвует в процессе эксцизионного восстановления оснований ДНК |
| Эндо Р[16] | Клетки HeLa | Специально для сайтов GC |
Мутации
Пигментная ксеродермия это редкое аутосомно-рецессивное заболевание, вызванное дефектной УФ-специфической эндонуклеазой. Пациенты с мутациями не могут восстановить повреждение ДНК, вызванное солнечным светом.[17]
Серповидно-клеточная анемия - это заболевание, вызванное точечной мутацией. Последовательность, измененная мутацией, устраняет сайт узнавания для эндонуклеазы рестрикции MstII, которая распознает нуклеотидную последовательность.[18]
Мутации эндонуклеазы сплайсинга тРНК вызывают понтоцеребеллярную гипоплазию. Понтоцеребеллярная гипоплазия (PCH) представляет собой группу нейродегенеративных аутосомно-рецессивных заболеваний, которые вызваны мутациями в трех из четырех различных субъединиц эндонуклеазного комплекса, связывающего тРНК.[19]
Смотрите также
Рекомендации
- ^ «Свойства экзонуклеаз и эндонуклеаз». New England BioLabs. 2017. Получено 21 мая, 2017.
- ^ Слор, Ханох (14 апреля 1975 г.). «Различие между экзонуклеазами и эндонуклеазами, а также между гаплотомными и диплотомными эндонуклеазами с использованием покрытых 3H-ДНК лунок пластиковых депрессионных планшетов в качестве субстрата». Исследования нуклеиновых кислот. 2 (6): 897–903. Дои:10.1093 / nar / 2.6.897. ЧВК 343476. PMID 167356.
- ^ Стивен Т. Килпатрик; Джоселин Э. Кребс; Левин, Бенджамин; Гольдштейн, Эллиотт (2011). Гены Левина X. Бостон: Джонс и Бартлетт. ISBN 978-0-7637-6632-0.
- ^ а б c d е ж Кокс М., Нельсон Д.Р., Ленингер А.Л. (2005). Принципы биохимии Ленингера. Сан-Франциско: W.H. Фримен. стр.952. ISBN 978-0-7167-4339-2.
- ^ Саймон М (2010). Новые вычисления: акцент на биоинформатике. Нью-Йорк: Спрингер. п. 437. ISBN 978-1441919632.
- ^ а б c Смит, Х.о .; Натанс, Д. (15 декабря 1973 г.). «Предлагаемая номенклатура для бактериальных систем модификации и рестрикции хозяев и их ферментов». Журнал молекулярной биологии. 81 (3): 419–23. Дои:10.1016/0022-2836(73)90152-6. PMID 4588280.
- ^ Рубин, РА; Модрич, П. (25 октября 1977 г.). «EcoRI метилаза». Журнал биологической химии. 252 (20): 7265–72. PMID 332688.
- ^ Лосик Р., Уотсон Дж. Д., Бейкер Т. А., Белл С., Ганн С., Левин М. В. (2008). Молекулярная биология гена. Сан-Франциско: Пирсон / Бенджамин Каммингс. ISBN 978-0-8053-9592-1.
- ^ Элленбергер Т., Фридберг ЕС, Уокер Г.С., Вольфрам С., Вуд Р.Дж., Шульц Р. (2006). Ремонт ДНК и мутагенез. Вашингтон, округ Колумбия: ASM Press. ISBN 978-1-55581-319-2.
- ^ Альбертс Б. (2002). Молекулярная биология клетки. Нью-Йорк: Наука Гарланд. ISBN 978-0-8153-3218-3.
- ^ Нишино Т., Морикава К. (декабрь 2002 г.). «Структура и функция нуклеаз в репарации ДНК: форма, захват и лезвие ножниц для ДНК». Онкоген. 21 (58): 9022–32. Дои:10.1038 / sj.onc.1206135. PMID 12483517.
- ^ Таня А. Бейкер; Корнберг, Артур (2005). Репликация ДНК. Университетская наука. ISBN 978-1-891389-44-3.
- ^ Wei, CF; Alianell, GA; Bencen, GH; Грей HB, младший (25 ноября 1983 г.). «Выделение и сравнение двух молекулярных видов нуклеазы BAL 31 из Alteromonas espejiana с различными кинетическими свойствами». Журнал биологической химии. 258 (22): 13506–12. PMID 6643438.
- ^ Linn, S; Lehman, IR (10 июня 1966 г.). «Эндонуклеаза митохондрий Neurospora crassa». Журнал биологической химии. 241 (11): 2694–9. PMID 4287861.
- ^ Холломан, WK; Холлидей, Р. (10 декабря 1973 г.). «Исследования нуклеазы из Ustilago maydis. I. Очистка, свойства и участие в рекомбинации фермента». Журнал биологической химии. 248 (23): 8107–13. PMID 4201782.
- ^ Готтлиб, Дж; Muzyczka, N (5 июля 1990 г.). «Очистка и характеристика эндонуклеазы HeLa R. G-специфической эндонуклеазы млекопитающих». Журнал биологической химии. 265 (19): 10836–41. PMID 2358441.
- ^ Медицинская биохимия вкратце. Нью-Йорк: Вили. 2012 г. ISBN 978-0-470-65451-4.
- ^ Ферриер Д.Р., Чампе П.С., Харви Р.П. (2008). Биохимия. Филадельфия: Уолтерс Клувер / Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-6960-0.
- ^ Бадде Б.С., Намавар Й., Барт П.Г., Опрос-БТ, Нюрнберг Дж., Беккер К., ван Рейссен Ф., Ветерман М.А., Флюитер К., те Бик Е.Т., Ароника Е., ван дер Кнаап М.С., Хоне В., Толиат М.Р., Ворон Ю.Дж. , Steinling M, Voit T, Roelenso F, Brussel W, Brockmann K, Kyllerman M, Boltshauser E, Hammersen G, Willemsen M, Basel-Vanagaite L, Krägeloh-Mann I, de Vries LS, Sztriha L, Muntoni F, Ferrie CD , Battini R, Hennekam RC, Grillo E, Beemer FA, Stoets LM, Wollnik B, Nürnberg P, Baas F (сентябрь 2008 г.). «Мутации эндонуклеазы сплайсинга тРНК вызывают понтоцеребеллярную гипоплазию». Nat. Genet. 40 (9): 1113–8. Дои:10,1038 / нг.204. PMID 18711368.