Сфингомиелин фосфодиэстераза - Sphingomyelin phosphodiesterase
| Сфингомиелин фосфодиэстераза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Кристаллическая структура сфингомиелиназы из Bacillus cereus.[1] | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 3.1.4.12 | ||||||||
| Количество CAS | 9031-54-3 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
Сфингомиелин фосфодиэстераза (EC 3.1.4.12, также известный как нейтральная сфингомиелиназа, сфингомиелиназа, или же SMase) это гидролаза фермент, который участвует в сфинголипид реакции обмена веществ. SMase является членом ДНКаза I суперсемейство ферментов и отвечает за нарушение сфингомиелин (SM) вниз в фосфохолин и керамид. Активация SMase была предложена в качестве основного пути производства церамида в ответ на клеточные стрессы.[2]
Семейство сфингомиелиназ
Выявлено пять типов SMase. Они классифицируются в соответствии с их катионной зависимостью и оптимумом действия pH и включают:
- Лизосомальный кислота SMase
- Секретируемый цинк-зависимый кислота SMase
- Магний-зависимый нейтральный SMase
- Независимый от магния нейтральный SMase
- Щелочная SMase
Из них лизосомальная кислая SMase и магний-зависимая нейтральная SMase считаются основными кандидатами на продукцию церамида в клеточной реакции на стресс.
Нейтральная сфингомиелиназа
Активность нейтральной сфингомиелиназы (N-SMase) была впервые описана в фибробластах пациентов с Болезнь Ниманна-Пика - лизосомная болезнь накопления, характеризующаяся дефицитом кислой SMase.[3] Последующее исследование показало, что этот фермент был продуктом отдельного гена, имел оптимальный pH 7,4, зависел от Mg.2+ ионами для активности и особенно обогащен мозгом.[4] Однако более недавнее исследование головного мозга крупного рогатого скота показало, что существует множество изоформ N-SMase с разными биохимическими и хроматографическими свойствами.[5]
Главный прорыв произошел в середине 1980-х годов с клонированием первых N-SMases из Bacillus cereus и Золотистый стафилококк.[6][7] Использование последовательностей этих бактериальных сфингомиелиназ в поисках гомологии в конечном итоге привело к идентификации дрожжевых N-SMases ISC1 в почкующихся дрожжах. Saccharomyces cerevisiae[8] и ферменты N-SMase млекопитающих, nSMase1 и nSMase2.[9][10][11] Идентичность между SMase млекопитающих, дрожжей и бактерий очень низка - примерно 20% между nSMase2 и SMase B. cereus. Однако выравнивание последовательностей (см. Рисунок) указывает на ряд консервативных остатков во всем семействе, особенно в каталитической области ферментов.[12] Это привело к предположению об общем каталитическом механизме для семейства N-SMase.
Третий белок N-SMase, называемый nSMase3 - недавно был[когда? ] клонировано и охарактеризовано. nSMase3 имеет небольшое сходство последовательностей с nSMase1 или nSMase2. Однако, по-видимому, существует высокая степень эволюционной консервативности от низших организмов к высшим, что позволяет предположить, что они могут включать уникальную и отличную N-SMase. Высокая экспрессия nSMase3 в сердце и скелетных мышцах также предполагает потенциальную роль в работе сердца.[13]
Активный сайт

Решение кристаллической структуры нейтральной сфингомиелиназы из Listeria ivanovii и Bacillus cereus позволил более полно понять их ферментативный сайт. В активный сайт из B. cereus SMase состоит из остатков Asn -16, Glu -53, Жерех -195, Асн-197 и Его -296. Известно, что из них остатки Glu-53, Asp-195 и His-296 важны для активности. Относительная каталитическая активность SMase, когда ионы металлов связаны с активным центром, была изучена для ионов двухвалентных металлов Co2+, Mn2+, Mg2+, Ca2+, а Sr2+. Из этих пяти ионов металлов Co2+, Mn2+, и Mg2+ связанные с активным сайтом, приводят к высокой каталитической активности SMase. Ca2+ и Sr2+ связаны с активным сайтом, гораздо ниже каталитический активность SMase. Когда один Mg2+ ион или два Co2+ ионы связываются с активным центром, двойной гекса-согласованный геометрические результаты с двумя октаэдрическими бипирамидами для Co2+ и один восьмигранный бипирамида для Mg2+. Когда один Ca2+ ion связывается с активным центром, в результате получается гепта-скоординированная геометрия. Следовательно, предполагается, что различие в каталитической активности для ионов металлов связано с геометрическими различиями. Ко2+ и Mg2+, SMase имеет лучшую реактивность, когда два Co2+ ионы связаны с SMase; когда эти Co2+ ионы связаны, Glu-53 и His-296 каждый связывает один катион двухвалентного металла. Эти катионы окружены мостиковыми молекулами воды и действуют как Кислоты Льюиса.[1]
Механизм
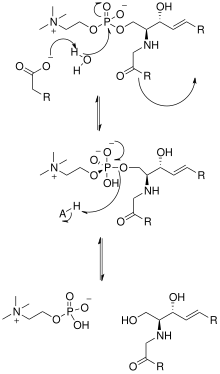
Решение кристаллической структуры нейтральной сфингомиелиназы из Listeria ivanovii и Bacillus cereus также пролил свет на их каталитические механизмы. Активный центр SMase содержит остатки Glu и His, каждый из которых связан с одним или двумя катионами двухвалентного металла, обычно Co2+, Mg2+, или Ca2+ для оптимальной производительности. Эти два катиона участвуют в катализе, привлекая SM к активному центру SMase. Двухвалентный катион, связанный с остатком Glu, взаимодействует с амидокислородом и сложный эфир -кислород между C1 и фосфат группа СМ; Остаток Asn и катион двухвалентного металла, связанный с остатком His, связываются с атомами кислорода фосфатной группы SM. Это стабилизирует отрицательный заряд фосфатной группы. Катион металла, связанный с остатком His и боковыми цепями Asp и Asn, снижает pKa значение одной из мостиковых молекул воды, таким образом активируя молекулу воды. Эта молекула воды затем действует как нуклеофил и атакует фосфатную группу SM, создавая пятивалентный атом фосфора, отрицательный заряд которого стабилизируется катионами двухвалентных металлов. Затем фосфат восстанавливает свой четырехгранный соответствие и результаты в продуктах керамид и фосфохолин.[1] Однако в настоящее время[когда? ] неясно, является ли механизм действия кислой сфингомиелиназы таким же из-за отсутствия кристаллической структуры.
Рекомендации
- ^ а б c PDB: 2ddt; Аго Х., Ода М., Такахаши М., Цуге Х., Очи С., Катунума Н., Мияно М., Сакураи Дж. (Июнь 2006 г.). «Структурные основы активности сфингомиелин фосфодиэстеразы в нейтральной сфингомиелиназе из Bacillus cereus». J. Biol. Chem. 281 (23): 16157–67. Дои:10.1074 / jbc.M601089200. PMID 16595670.
- ^ Ханнун Ю.А., Обейд Л.М. (июль 2002 г.). «Церамид-центрическая вселенная липидно-опосредованной регуляции клеток: стрессовые столкновения липидного типа». J. Biol. Chem. 277 (29): 25847–50. Дои:10.1074 / jbc.R200008200. PMID 12011103.
- ^ Шнайдер ПБ, Кеннеди EP (май 1967). «Сфингомиелиназа в нормальной селезенке человека и в селезенке субъектов с болезнью Ниманна-Пика». J. Lipid Res. 8 (3): 202–9. PMID 4962590.
- ^ Рао Б.Г., Спенс М.В. (сентябрь 1976 г.). «Активность сфингомиелиназы при pH 7,4 в мозге человека и сравнение с активностью при pH 5,0». J. Lipid Res. 17 (5): 506–15. PMID 9463.
- ^ Юнг СИ, Сух Дж. Х., Пак Х. Дж., Юнг К. М., Ким М. Ю., На Д. С., Ким Д. К. (сентябрь 2000 г.). «Идентификация множественных форм мембран-ассоциированной нейтральной сфингомиелиназы в головном мозге крупного рогатого скота». J. Neurochem. 75 (3): 1004–14. Дои:10.1046 / j.1471-4159.2000.0751004.x. PMID 10936181. S2CID 46397368.
- ^ Коулман, округ Колумбия, Арбутнотт, JP, Помрой, Х.М., Биркбек, TH (декабрь 1986). «Клонирование и экспрессия в Escherichia coli и Staphylococcus aureus детерминанты бета-лизина из Staphylococcus aureus: доказательства того, что бактериофаговое преобразование активности бета-лизина вызвано инсерционной инактивацией детерминанты бета-лизина». Microb. Патог. 1 (6): 549–64. Дои:10.1016/0882-4010(86)90040-9. PMID 3334158.
- ^ Ямада А., Цукагоши Н., Удака С., Сасаки Т., Макино С., Накамура С., Литтл К., Томита М., Икэдзава Х (август 1988 г.). «Нуклеотидная последовательность и экспрессия в Escherichia coli гена, кодирующего сфингомиелиназу Bacillus cereus». Евро. J. Biochem. 175 (2): 213–20. Дои:10.1111 / j.1432-1033.1988.tb14186.x. PMID 2841128.
- ^ Савай Х., Окамото Ю., Луберто С., Мао С., Белавска А., Домае Н., Ханнун Ю.А. (декабрь 2000 г.). «Идентификация ISC1 (YER019w) как инозитолфосфосфинголипидфосфолипаза C в Saccharomyces cerevisiae». J. Biol. Chem. 275 (50): 39793–8. Дои:10.1074 / jbc.M007721200. PMID 11006294.
- ^ Трипати, К. (2015). «Роль инозитолфосфинголипидфосфолипазы C1, дрожжевого гомолога нейтральных сфингомиелиназ в ответе на повреждение ДНК и заболеваниях». Журнал липидов. 2015: 161392. Дои:10.1155/2015/161392. ЧВК 4544949. PMID 26346287.
- ^ Томюк С., Хофманн К., Никс М., Зумбансен М., Стоффель В. (март 1998 г.). "Клонированная нейтральная сфингомиелиназа млекопитающих: функции в передаче сигналов сфинголипидов?". Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 95 (7): 3638–43. Bibcode:1998PNAS ... 95.3638T. Дои:10.1073 / пнас.95.7.3638. ЧВК 19888. PMID 9520418.
- ^ Томюк С., Зумбансен М., Стоффель В. (февраль 2000 г.). «Характеристика и субклеточная локализация магний-зависимой нейтральной сфингомиелиназы человека и мыши». J. Biol. Chem. 275 (8): 5710–7. Дои:10.1074 / jbc.275.8.5710. PMID 10681556.
- ^ Кларк CJ, Snook CF, Tani M, Matmati N, Marchesini N, Hannun YA (сентябрь 2006 г.). «Расширенная семья нейтральных сфингомиелиназ». Биохимия. 45 (38): 11247–56. Дои:10.1021 / bi061307z. PMID 16981685.
- ^ Krut O, Wiegmann K, Kashkar H, Yazdanpanah B, Krönke M (май 2006 г.). «Новая нейтральная сфингомиелиназа-3 млекопитающих, чувствительная к фактору некроза опухоли, представляет собой белок, заякоренный в С-хвосте». J. Biol. Chem. 281 (19): 13784–93. Дои:10.1074 / jbc.M511306200. PMID 16517606.
дальнейшее чтение
внешняя ссылка
- Сфингомиелин + фосфодиэстераза в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)