RecBCD - RecBCD
| Экзодезоксирибонуклеаза V | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 3.1.11.5 | ||||||||
| Количество CAS | 37350-26-8 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
RecBCD (EC 3.1.11.5, Экзонуклеаза V, Экзонуклеаза V Escherichia coli, Экзонуклеаза V E. coli, ген recBC эндофермент, RecBC дезоксирибонуклеаза, ген recBC ДНКаза, ферменты ген recBCD) является фермент из Кишечная палочка бактерия, которая инициирует рекомбинационная репарация от потенциально смертельного двухрядные разрывы в ДНК что может быть результатом ионизирующее излучение, ошибки репликации, эндонуклеазы, окислительное повреждение и множество других факторов.[2][3] Фермент RecBCD является одновременно геликаза который раскручивает или разделяет нити ДНК, и нуклеаза это делает одноцепочечные разрывы в ДНК.[1]
Структура
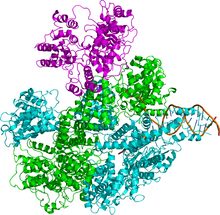
Ферментный комплекс состоит из трех разных субъединиц, называемых RecB, RecC и RecD, и поэтому комплекс называется RecBCD (рис. 1). До открытия recD ген,[4] фермент был известен как «RecBC». Каждая субъединица кодируется отдельным геном:
| ген | цепь | белок | функция |
|---|---|---|---|
| RecB | бета | P08394 | 3'-5 'геликаза, нуклеаза |
| RecC | гамма | P07648 | Узнает Чи (cРоссовер часотспот яnstigator) |
| RecD | альфа | P04993 | 5'-3 'геликаза |
Функция

Субъединицы RecD и RecB представляют собой геликазы, т.е., энергозависимые молекулярные моторы, которые раскручивают ДНК (или РНК в случае других белков). Субъединица RecB, кроме того, выполняет нуклеазную функцию.[5] Наконец, фермент RecBCD (возможно, субъединица RecC) распознает определенную последовательность в ДНК, 5' -GCTGGTGG-3', известный как Чи (иногда обозначается греческой буквой χ).
RecBCD необычен среди геликасов, потому что он имеет две геликасы, которые перемещаются с разной скоростью.[6] и потому, что он может распознавать и изменять последовательность ДНК Чи.[7][8] RecBCD активно связывает конец линейной двухцепочечной (ds) ДНК. Хеликаза RecD перемещается по цепи с 5 'концом, на котором фермент инициирует раскручивание, и RecB по цепи с 3' концом. RecB работает медленнее, чем RecD, так что одноцепочечная (ss) петля ДНК накапливается впереди RecB (рисунок 2). При этом образуются структуры ДНК с двумя ss-хвостами (более короткий хвост с 3’-концом и более длинный хвост с 5 ’конца) и одной ss-петлей (на цепи с 3’-концом), наблюдаемые с помощью электронной микроскопии.[9] SS-хвосты можно отжечь, чтобы получить второй ss-контур, дополнительный к первому; такие двухпетлевые структуры первоначально назывались «кроличьи уши».
Механизм действия

Во время раскрутки нуклеаза в RecB может действовать по-разному в зависимости от условий реакции, а именно от соотношения концентраций Mg2+ ионы и АТФ. (1) Если АТФ в избытке, фермент просто надрывает цепь Chi (цепь с начальным 3 'концом) (рис. 2).[10][11] Раскручивание продолжается и образует 3-й хвост с Чи около его конца. Этот хвост может быть связан с белком RecA, который способствует обмену цепей с интактным гомологичным дуплексом ДНК.[12] Когда RecBCD достигает конца ДНК, все три субъединицы разбираются, и фермент остается неактивным в течение часа или более;[13] молекула RecBCD, действующая на Chi, не атакует другую молекулу ДНК. (2) Если Mg2+ ионы в избытке, RecBCD расщепляет обе нити ДНК эндонуклеолитически, хотя 5'-хвост расщепляется реже (рис. 3).[14] Когда RecBCD встречает сайт Чи на 3-концевой цепи, паузы при раскручивании и переваривание 3-футового хвоста уменьшаются.[15] Когда RecBCD возобновляет раскручивание, он теперь отсекает противоположную нить (т.е., 5 'хвост)[16][17] и загружает белок RecA на 3’-концевую цепь.[12] После завершения реакции на одной молекуле ДНК фермент быстро атакует вторую ДНК, на которой происходят те же реакции, что и на первой ДНК.
Хотя ни одна из реакций не была подтверждена анализом внутриклеточной ДНК, из-за временного характера промежуточных продуктов реакции, генетические данные показывают, что первая реакция более близка к реакции в клетках.[2] Например, на активность Chi влияют нуклеотиды на его 3'-стороне, как в клетках, так и в реакциях с избытком АТФ, но не с Mg.2+ в избытке [PMID 27401752, 27330137]. Мутанты RecBCD, лишенные детектируемой экзонуклеазной активности, сохраняют высокую активность горячих точек Chi в клетках и надрывают Chi за пределами клеток.[18] Сайт Chi на одной молекуле ДНК в клетках снижает или устраняет активность Chi на другой ДНК, возможно, отражая Chi-зависимую разборку RecBCD, наблюдаемую in vitro в условиях избытка АТФ и расщепления ДНК на Chi.[19][20]
В обоих условиях реакции 3'-цепь остается нетронутой после Chi. В RecA Затем белок активно загружается на 3'-хвост с помощью RecBCD.[12] В какой-то неопределенный момент RecBCD отделяется от ДНК, хотя RecBCD может раскрутить по крайней мере 60 т.п.н. ДНК без отрыва. RecA инициирует обмен нити ДНК, с которой он связан, с идентичной или почти идентичной нитью в интактном дуплексе ДНК; этот обмен цепями генерирует совместную молекулу ДНК, такую как D-петля (рис. 2). Считается, что объединенная молекула ДНК разделяется либо путем репликации, примированной вторгающейся цепью с 3’-концом, содержащей Chi, либо путем расщепления D-петли и образования соединения Холлидея. В Холлидей Джанкшн можно преобразовать в линейную ДНК RuvABC сложный или диссоциированный RecG белок. Каждое из этих событий может генерировать неповрежденную ДНК с новыми комбинациями генетических маркеров, по которым родительские ДНК могут отличаться. Этот процесс, гомологичная рекомбинация, завершает репарацию двухцепочечного разрыва ДНК.
Приложения
RecBCD - это модельный фермент для использования флуоресценция одной молекулы как экспериментальный метод, используемый для лучшего понимания функции взаимодействий белок-ДНК.[21] Фермент также полезен для удаления линейной ДНК, одно- или двухцепочечной, из препаратов кольцевой двухцепочечной ДНК, поскольку для его активности требуется конец ДНК.
использованная литература
- ^ а б Синглтон М. Р., Диллингем М. С., Годье М., Ковальчиковски С. К., Вигли Д. Б. (ноябрь 2004 г.). «Кристаллическая структура фермента RecBCD раскрывает машину для обработки разрывов ДНК». Природа. 432 (7014): 187–93. Bibcode:2004Натура.432..187С. Дои:10.1038 / природа02988. PMID 15538360.
- ^ а б Смит, Г. Р. (июнь 2012 г.). «Как фермент RecBCD и хи способствуют восстановлению разрывов ДНК и рекомбинации: взгляд молекулярных биологов». Микробиол Мол Биол Рев. 76 (2): 217–28. Дои:10.1128 / MMBR.05026-11. ЧВК 3372252. PMID 22688812.
- ^ Шпионы М, Ковальчиковски SC (2003). «Гомологичная рекомбинация по путям RecBCD и RecF». В Хиггинсе P (ред.). Бактериальные хромосомы. Вашингтон, округ Колумбия: ASM Press. С. 389–403. ISBN 1-55581-232-5.
- ^ Амундсен С.К., Тейлор А.Ф., Чаудхури А.М., Смит Г.Р. (август 1986 г.). "recD: ген важной третьей субъединицы экзонуклеазы V. " Proc Natl Acad Sci U S A. 83 (15): 5558–62. Bibcode:1986PNAS ... 83.5558A. Дои:10.1073 / pnas.83.15.5558. ЧВК 386327. PMID 3526335.
- ^ Ю. М., Суая Дж., Джулин Д. А. (февраль 1998 г.). «С-концевой домен 30 кДа белка RecB имеет решающее значение для нуклеазной активности, но не для активности геликазы фермента RecBCD из кишечная палочка". Proc Natl Acad Sci U S A. 95 (3): 981–6. Дои:10.1073 / пнас.95.3.981. ЧВК 18645. PMID 9448271.
- ^ Тейлор А.Ф., Смит Г.Р. (июнь 2003 г.). «Фермент RecBCD - это ДНК-геликаза с быстрым и медленным двигателями противоположной полярности». Природа. 423 (6942): 889–93. Bibcode:2003Натура 423..889Т. Дои:10.1038 / природа01674. PMID 12815437.
- ^ Тейлор А.Ф., Смит Г.Р. (июнь 1992 г.). «Фермент RecBCD изменяется при разрезании ДНК в горячей точке рекомбинации Чи». Proc Natl Acad Sci U S A. 89 (12): 5226–30. Bibcode:1992ПНАС ... 89.5226Т. Дои:10.1073 / пнас.89.12.5226. ЧВК 49264. PMID 1535156.
- ^ Амундсен СК, Тейлор А.Ф., Редди М., Смит Г.Р. (декабрь 2007 г.). «Межсубъединичная передача сигналов в ферменте RecBCD, сложной белковой машине, регулируемой горячими точками Ци». Genes Dev. 21 (24): 3296–307. Дои:10.1101 / gad.1605807. ЧВК 2113030. PMID 18079176.
- ^ Тейлор А., Смит Г. Р. (ноябрь 1980 г.). «Размотка и перемотка ДНК ферментом RecBC». Ячейка. 22 (2 Pt 2): 447–57. Дои:10.1016/0092-8674(80)90355-4. PMID 6256081.
- ^ Ponticelli, A. S .; Schultz, D. W .; Тейлор, А. Ф .; Смит, Г. Р. (май 1985 г.). «Хи-зависимое расщепление цепи ДНК ферментом RecBC». Ячейка. 41 (1): 145–151. Дои:10.1016/0092-8674(85)90069-8. PMID 3888404.
- ^ Тейлор А.Ф., Шульц Д.В., Понтичелли А.С., Смит Г.Р. (май 1985 г.). «Расщепление фермента RecBC на сайтах Chi во время раскручивания ДНК: расположение и ориентация разрезания». Ячейка. 41 (1): 153–63. Дои:10.1016/0092-8674(85)90070-4. PMID 3888405.
- ^ а б c Андерсон Д.Г., Ковальчиковски С.К. (июль 1997 г.). «Транслоцирующий фермент RecBCD стимулирует рекомбинацию, направляя белок RecA на оцДНК посредством Chi». Ячейка. 90 (1): 77–86. Дои:10.1016 / S0092-8674 (00) 80315-3. PMID 9230304.
- ^ Тейлор А.Ф., Смит Г.Р. (апрель 1999 г.). «Регулирование гомологичной рекомбинации: Chi инактивирует фермент RecBCD путем разборки трех субъединиц». Genes Dev. 13 (7): 890–900. Дои:10.1101 / gad.13.7.890. ЧВК 316601. PMID 10197988.
- ^ Dixon DA, Kowalczykowski SC (апрель 1993 г.). «Горячая точка рекомбинации Chi - это регуляторная последовательность, которая действует, ослабляя нуклеазную активность Кишечная палочка Фермент RecBCD ». Ячейка. 73 (1): 87–96. Дои:10.1016 / 0092-8674 (93) 90162-Дж. PMID 8384931.
- ^ Шпионы М., Амитани И., Баскин Р.Дж., Ковальчиковски С.К. (ноябрь 2007 г.). «Фермент RecBCD переключает двигательные субъединицы свинца в ответ на распознавание Чи». Ячейка. 131 (4): 694–705. Дои:10.1016 / j.cell.2007.09.023. ЧВК 2151923. PMID 18022364.
- ^ Тейлор А.Ф., Смит Г.Р. (октябрь 1995 г.). «Специфичность цепей ДНК в сайтах Chi ферментом RecBCD. Модуляция уровнями АТФ и магния». J Biol Chem. 270 (41): 24459–67. Дои:10.1074 / jbc.270.41.24459. PMID 7592661.
- ^ Андерсон Д.Г., Ковальчиковски С.К. (март 1997 г.). «Горячая точка рекомбинации chi является регуляторным элементом, который переключает полярность деградации ДНК ферментом RecBCD». Genes Dev. 11 (5): 571–81. Дои:10.1101 / gad.11.5.571. PMID 9119222.
- ^ Амундсен СК, Смит Г.Р. (январь 2007 г.). "Активность точек доступа Chi в кишечная палочка без активности экзонуклеазы RecBCD: последствия для механизма рекомбинации ». Генетика. 175 (1): 41–54. Дои:10.1534 / genetics.106.065524. ЧВК 1774988. PMID 17110484.
- ^ Кеппен А., Кробич С., Томс Б., Ваккернагель В. (июль 1995 г.). "Взаимодействие с горячей точкой рекомбинации Chi in vivo превращает фермент RecBCD в кишечная палочка в Chi-независимую рекомбиназу путем инактивации субъединицы RecD ». Proc Natl Acad Sci U S A. 92 (14): 6249–53. Bibcode:1995PNAS ... 92.6249K. Дои:10.1073 / пнас.92.14.6249. ЧВК 41495. PMID 7541534.
- ^ Майерс Р.С., Кузьминов А., Шталь Ф.В. (июль 1995 г.). "Горячая точка рекомбинации Chi активирует рекомбинацию RecBCD путем преобразования кишечная палочка к фенокопии с мутантом recD ". Proc Natl Acad Sci U S A. 92 (14): 6244–8. Bibcode:1995ПНАС ... 92.6244М. Дои:10.1073 / пнас.92.14.6244. ЧВК 41494. PMID 7603978.
- ^ Бьянко П.Р., Брюер Л.Р., Корцетт М., Балхорн Р., Йе Й, Ковальчиковски С.К., Баскин Р.Дж. (январь 2001 г.). «Процессивная транслокация и раскручивание ДНК отдельными молекулами фермента RecBCD». Природа. 409 (6818): 374–8. Bibcode:2001Натура.409..374Б. Дои:10.1038/35053131. PMID 11201750.
внешние ссылки
- Экзодезоксирибонуклеаза + V в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- Экзодезоксирибонуклеаза + V, + E + coli в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- EC 3.1.11.5